
Иванова В.С., Трошкина Н.Н. Контроль за развитием обыкновенного фазана // Искусственное разведение фазанов. Сборник научных трудов ЦНИЛ Главохоты РСФСР. Москва, 1983. С. 128-145.
Молодняк фазанов, выращиваемый на дичефермах и предназначенный для выпуска в угодья, должен быть не только жизнеспособным и крепким, но и сохранять экстерьерные показатели и повадки диких сородичей, которые до сих пор не до конца определены.
Данных по постэмбриональному развитию выводковых птиц, в том числе и фазанов, относительно мало, поскольку их молодняк очень подвижен сразу же после вылупления. Все исследования проведены либо на одомашненных формах, либо на диких видах, размножавшихся в неволе (Шмальгаузен, 1927, 1931, 1935 а, б, в; Ларионов и др., 1936; О.Хейнрот (Heinroth), 1924-1932; Кузнецов, 1972; Курскова, 1966; Э.Надь (Hady), 1971).
Данные по постэмбриогенезу подвидов обыкновенного фазанаограничиваются, как правило, констатацией живой массы взрослых птицили некоторыми морфометрическими показателями (Птицы Советского Союза, 1952; Определитель птиц фауны СССР, 1980; Кузьмина, 1964, 1977; Габузов и др. 1976 и др.).
Цель наших исследований - определение динамики весового роста, некоторых морфологических признаков и биохимических показателей в постэмбриогенезе у охотничьего, северокавказского и маньчжурского фазанов; разработка и предложение методов проведения прижизненного контроля за развитием молодняка. Материал собран в 1974-1982 гг. в дичепитомнике ЦНИЛ Главохоты РСФСР и в существующих в СССР фазанариях.
Фазанят тщательно сортировали уже с суточного возраста. Для эксперимента отбирали только здоровых птенцов: подвижных, активно реагирующих на звук, с мягким животом, закрытой пуповиной без следов кровотечений, чистой клоакой, рыхлым, ровным хорошо пигментированным, полностью прикрывающим пуповину пухом, симметричным клювом, крепкими ногами, с прямыми пальцами, плотно прилегающими ктелу крыльями. Масса тела зависела от принадлежности фазаненка к тому или иному подвиду (маньчжурский - 18-19 г, северокавказский - 21-22 г, охотничий - 22-23 г). Слабых фазанят или с пороками (чрезмерно высокий или чрезмерно низкий вес, слипшийся пух, плохо закрывающийся или искривленный клюв, искривленная шея, незатянутая пуповина, искривленные ноги или пальцы и т.д.) в эксперименте не использовали.
Наиболее существенным признаком для оценки фазанов была масса тела, которая меняясь с возрастом птиц, является показателем состояния организма. Птиц взвешивали индивидуально сначала на электрических весах ВЛК-500 с точностью до 1 г, затем, когда их масса превышала 500 г, - на торговых весах с точностью взвешивания до 5 г. Наблюдения за изменением массы тела трех форм фазанов в первые два месяца постэмбриогенеза проводили в следующих возрастах: суточном, 5, 10, 30, 30, 60-дневном. Массу взрослых птиц (самцов и самок отдельно) определяли весной (табл. 1). Из приведенных материалов видно, что у суточных птиц наименьшей массой обладали маньчжурские фазанята. Они легче охотничьих (на 3,6 г) исеверокавказских (на 3,5 г). В дальнейшем эти различия уменьшались и к 20-дневному возрасту масса тела у фазанят всех подвидов становилась одинаковой. К месячному возрасту картина опять менялась: самыми легкими были северокавказские фазанята, затем маньчжурские (на 7 г тяжелее) и, наконец, охотничьи (на 13 г тяжелее). У двухмесячных фазанов сохранялось такое же распределение: северокавказские на 8 г легче маньчжурских и на 11 г легче охотничьих. Эта тенденция, отмечалась и у взрослых птиц. Масса тела у северокавказских фазанов меньше, чем у маньчжурских (у самцов на 57 г, у самок - на 2г) и охотничьих фазанов (у самцов на 75 г, у самок - на 12,6 г).
Таблица 1
Показатели массы фазанов, выращенных в искусственных условиях (г)
|
Возраст (дней) |
Форма |
||
|
Маньчжурский |
Северокавказский |
Охотничий |
|
|
Вывод |
18,3±0,09 |
21,8±0,17 |
21,9±0,15 |
|
5 |
24,5±0,33 |
26,1±0,40 |
25,6±0,35 |
|
10 |
39,1±0,69 |
40,5±0,79 |
41,9±0,89 |
|
20 |
86,3±1,60 |
86,1±1,54 |
86,6±1,90 |
|
30 |
147,5±2,60 |
140,3±3,60 |
153,2±10,50 |
|
60 |
375,6±18,30 |
367,8±15,60 |
378,6±10,80 |
|
Взрослые: самцы |
1308,3±19,60 |
1251,4±18,53 |
1326,4±17,91 |
|
самки |
1000,7±17,30 |
1002,7±20,04 |
1015,3±21,30 |
К качественным показателям относится скорость роста, которая хорошо наследуется и отражает видовую особенность птиц.Для установления скорости роста производили индивидуальные взвешивания и измерения основных статей телосложения на значительном поголовье птиц.
О скорости роста судят как по абсолютной величине привеса, так и по относительному привесу, характеризующему интенсивность роста. Абсолютный привес определяют по изменению веса птицы (или отдельного промера) за известный промежуток времени по формуле:
V=V2-V1
где V1 - вес в начале периода (г); V2 - вес в конце периода (г). Так, если маньчжурский фазаненок к 30-дневному возрасту весил 147 г, а к 60-дневному возрасту - 375 г, то абсолютный привес его за месяц составлял 228 г (375-147). Среднесуточный абсолютный привес вычисляют по формуле:
V/t=V2-V1/t2-t1
где t1 - возраст на начало периода (дней); t2 - возраст в конце периода, (дней).
В нашем примере абсолютный суточный привес был равен:
375-147/60-30=7,6 г
Однако для характеристики скорости роста вычисление только абсолютного привеса недостаточно, так как величина его с возрастом меняется. Большой привес на более поздних стадиях развития не служит показателем интенсивности росса, а является лишь результатом увеличения растущей массы. Кроме того, абсолютные показатели не могут быть использованы при сравнении скорости роста птиц различных подвидов и, особенно, видов. Поэтому введена величина относительного привеса, вычисляемая по формуле Броди:
R= V2-V1/0,5(V2+V1)t x100
В нашем примере относительный привес был равен:
375-147/0,5(375+147)х30х100=2,9 %
По приведенным формулам определяли интенсивность весового роста трех форм фазанов.
Изтаблицы 2 видно, что увеличение массы тела у фазанов происходило равномерно. Так, абсолютные среднесуточные привесы увеличивались непрерывно от 1 до 7 г и достигли максимального показателя (7 г) для всех 3-х форм фазанов к 2-месячному возрасту, изменения относительных среднесуточных привесов происходили по-иному. Наибольшая величина их наблюдалась в раннем возрасте птиц, а затем снижалась.
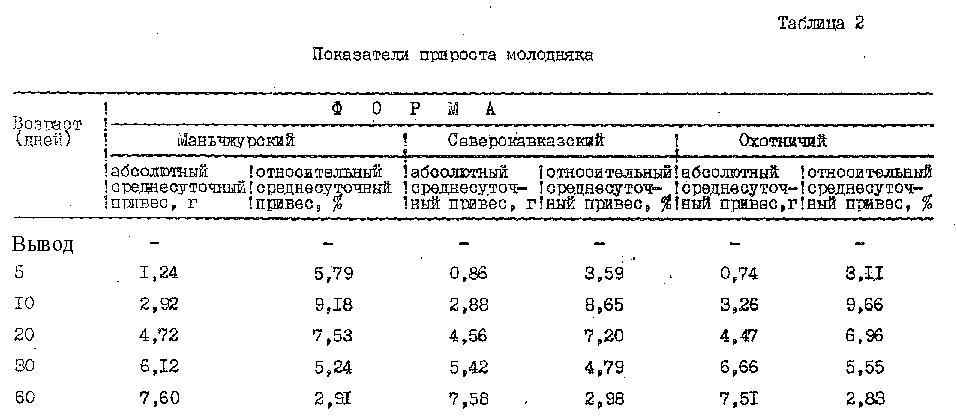
Сравнивая скорость роста изучаемых трех форм фазанов в течение 2-х месяцев постэмбриогенеза, можно отметить одинаковую интенсивность их весового роста, за исключением периода с 1 по 5 день. В это время маньчжурские фазаны росли несколько быстрее северокавказских и охотничьих фазанов (относительный среднесуточный привес их был на 2 %выше).
Для контроля за изменением телосложения фазанят проводили серию промеров: длина туловища - расстояние от плечелопаточного сочленения до основания копчиковой железы (сантиметровой лентой); обхват туловища - определяли сантметровой лентой у основания крыльев по линии, проходящей мимо заднего шейного позвонка и переднего конца киля; ширина таза - расстояние между выступами седалищных костей (штангенциркуль); длина головы - расстояние от мыщелка затылочной кости до вершины клюва (штангенциркуль); длина клюва - расстояние от основания клюва до его вершины или от ноздри до кончика клюва (штангенциркуль); длина предплечья - расстояние между крайними точками предплечья (штангенциркуль); длина кистевой части крыла - расстояние от кистевого сгиба до вершины самого длинного первостепенного махового пера (линейка); длина цевки - расстояние от точки соединения голени и плюсны вдоль нее до угла, образуемого третьим и четвертым пальцами (штангенциркуль); длина среднего пальца - расстояние от основания 1-ой фаланги до основания когтя (штангенциркуль); длина когтей - расстояние от основания каждого когтя до его вершины (штангенциркуль).
Кроме абсолютных цифр, мы вычисляли скорость роста по тем же формулам, по которым определяли скорость прироста массы тела (табл. 3, 4). Данные показывают, что величина прироста всех статей тела изменялась обратно пропорционально возрасту, в то время как абсолютные размеры с возрастом постепенно увеличивались. Наибольшая интенсивность роста характерна для первого месяца жизни всех трех форм фазанов. Нельзя также не отметить, что картина общего роста не однородна и разделяется на ряд отрезков, характеризующихся различным уровнем среднесуточного прироста или интенсивности роста. В первые 5 дней прирост еще невелик, в следующие 5 дней наблюдался самый интенсивный рост за весь период постэмбриогенеза. С 10 по 20 день интенсивность роста падала, а с 20 по 30 и, особенно, с 30 по 60 день величины относительных среднесуточных приростов всех статей тела были незначительны и снижались до 1 %. Следует отметить, что за 60 дней постэмбриогенеза с наибольшей интенсивностью росло крыло и предплечье, затем туловище (причем в длину интенсивнее, чем в ширину), далее клюв и цевка и медленнее всех росла голова.
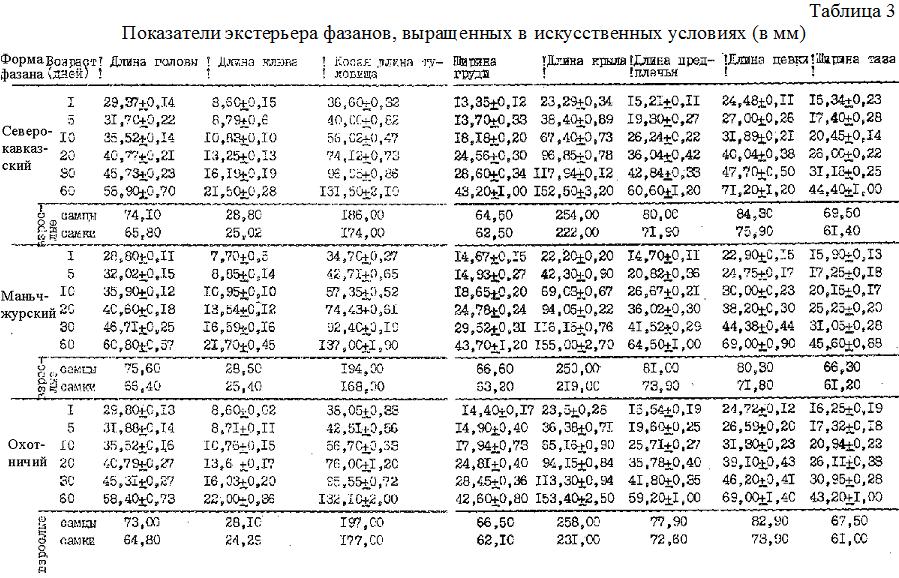
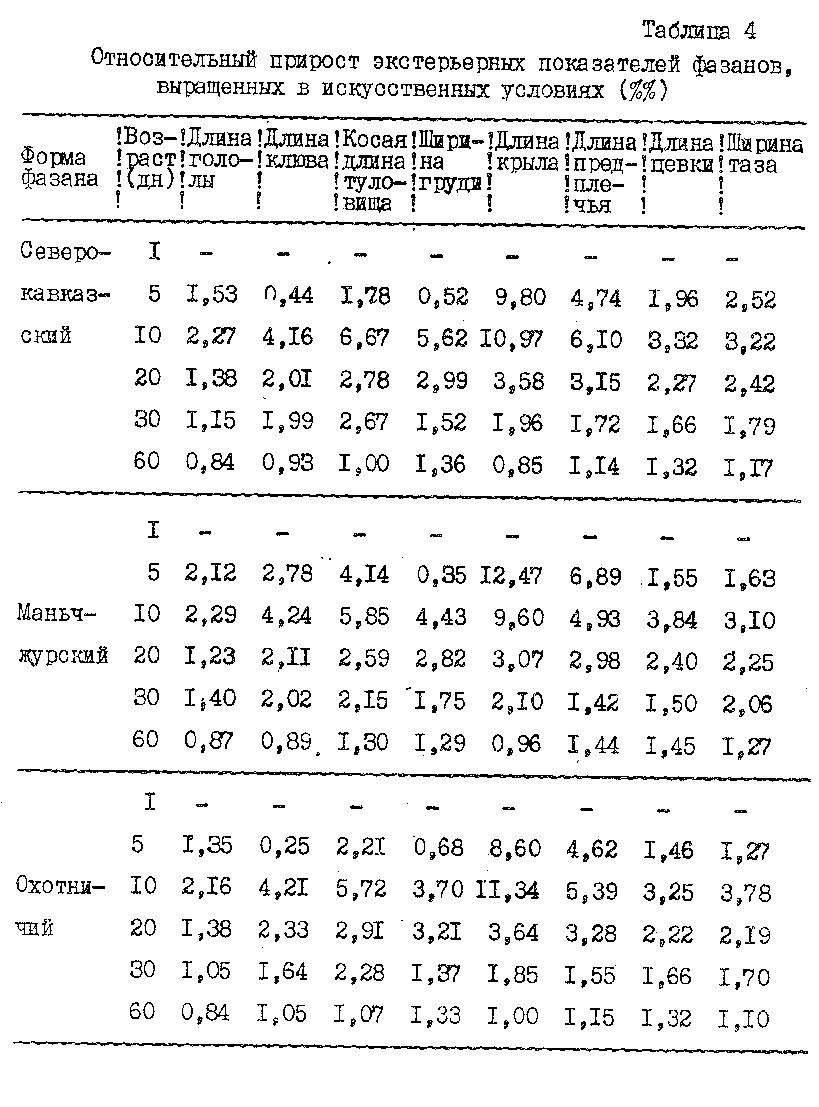
Из трех сравниваемых форм фазанов самыми мелкими (так же как и по живой массе) выводились маньчжурские, и самыми крупными - охотничьи фазанята; северокавказские же занимали среднее положение. За 20 дней постэмбриогенеза, имея более высокий темп роста большинства статей тела, маньчжурский фазан в конце 2-ой декады по размерам почти не отличался от северокавказского и охотничьего фазанов. А во взрослом состоянии у него даже появлялось преимущество в размерах некоторых статей тела, например, таких, как длина головы, клюва, предплечья и ширина груди. По длине крыла и цевки он уступал двум остальным формам фазанов. Охотничьи фазаны имели более длинное туловище и крыло, оперение крыла сохранилось у них в лучшем состоянии. Полученное преимущество в длине крыла у охотничьего фазана по сравнению с северокавказским и маньчжурским, очевидно, связано с более спокойным поведением при выращивании. Северокавказские и, особенно, маньчжурские фазаны, начиная с 30-дневного возраста, отличались пугливостью и дикостью, при малейшем беспокойстве резко взлетали вверх и оббивалимаховые перья крыла.
Северокавказский фазан по большинству промеров занимал промежуточное положение между маньчжурским и охотничьим, отличаясь лишь более развитой цевкой и тазом.
Таким образом, увеличение размеров тела фазанят происходит постоянно ипостепенно; величины приростов статей тела обратно пропорциональны возрасту фазанов. Наибольшая интенсивность роста у фазанов всех подвидов наблюдается в первый месяц жизни; суточные маньчжурские фазанята по размерам тела (и по массе) уступают северокавказским и охотничьим. Взрослые маньчжурские фазаны имеют преимущество в длине головы, клюва, предплечья и ширине груди. У охотничьих фазанов самое длинное туловище, а у северокавказских фазанов - более длинная цевка и широкий таз.
Развитие оперения и линька у отечественных подвидов обыкновенного фазана, разводимых в искусственных условиях, пока исследованы недостаточно. Изучение данного вопроса крайне необходимо, т.к. процесс развития оперения исмена перьев служит показателем физиологического состояния организма птицы. Так, например, на сельскохозяйственных птицах замечено, если помимо обычной ежегодной линьки в более или менее постоянные сроки, наступает ранняя, возникает внеочередная или начавшаяся линька затягивается (в результате чего яйцекладка снижается), это указывает на неправильное кормление, содержание или на заболевание птицы.
В наших опытах окрас пухового оперения у только что вылупившихся фазанят варьировал от светложелтого до оранжевого, на голове были три черные полоски, переходящие на спину. Спустя 11-18 часов длина маховых перьев первого порядка достигала уже 3-5 мм. По длине первичных маховых перьев у суточного молодняка сельскохозяйственных птиц судят о быстроте оперяемости. Определение оперяемости птенца в суточном возрасте требует некоторого навыка. После того как он обсохнет, необходимо осмотреть на ярком свету развернутое крыло, обратить внимание на длину маховых перьев на нижней стороне крыла.
В 10-12-дневном возрасте сложенные крылья уже полностью закрывали туловище фазаненка, плечи были покрыты кроющим пером, рулевые перья выходили из пеньков на 2-3 мм и появлялись пеньки второстепенных рулевых перьев. Развитие оперения способствовало возникновению качественно нового способа передвижения птенцов: если маленькие фазанята передвигались только по земле, то в этом возрасте они уже вспархивали на высоту 0,5-1 метр.
У 30-дневных фазанят полностью формировалось первое оперение, появился половой диморфизм - у самцов на груди и хвосте становилось заметным промежуточное перо. Эти красновато-бурые перья, в отличие от взрослого пера, не имели металлического отлива. Начиналась линька первого птенцового оперения - выпадали внутренние маховые и обламывались рулевые перья, первым выпадало 10-е перо. Маховые перья, как и у всех куриных, менялись от внутреннего края к наружному, то есть сохранялась несущая поверхность крыла и птица не лишалась способности к полету. Промежуток между выпадением соседних птенцовых маховых у фазанят исчисляется сутками, эта величина являлась показателем интенсивности линьки.
В 60-дневном возрасте заканчивался рост птенцового оперения. На груди, нижней половине шеи, спине появлялись пеньки взрослого оперения (у самцов красноватые перья с металлическим блеском; у охотничьего фазана на нежней трети спины появлялись буро-зеленые перья). Шла линька птенцовых и рост дефинитивных маховых и рулевых перьев. У птиц сменялись 4-5 (10, 9, 8, 7, 6 перья) первостепенных маховых перьев.
У фазанов, как у всех куриных птиц, наблюдалась постепенность смены ювенальных перьев на дефинитивные, когда на теле птенца одновременно находились перья обеих генераций.
Сохранность (жизнеспособность) птиц - количественный показатель, обусловливающий экономическую эффективность разведения фазанов в искусственных условиях, так как определяет выход продукции (количество голов) и влияет на ее себестоимость. При благополучном состоянии хозяйства по инфекционным и инвазионным заболеваниям и соблюдении всех правил кормления и содержания сохранность фазанят такова: за период выращивания с 1 по 10 день – 90 %; с 10 по 60 день – 80 %; с 60 по 280 день – 90 %. Следует заметить, что в нашем опыте на первые 80 дней выращивания приходился самый большой (13 %) процент падежа. Смертность фазанят росла к 5-7 дню выращивания, тогда же наблюдался ее максимум, а затем, к концу 2-ой декады, она снижалась до минимума. Гибель фазанят в этот критический период связана, по-видимому, с перестройкой работы кишечного тракта птиц.
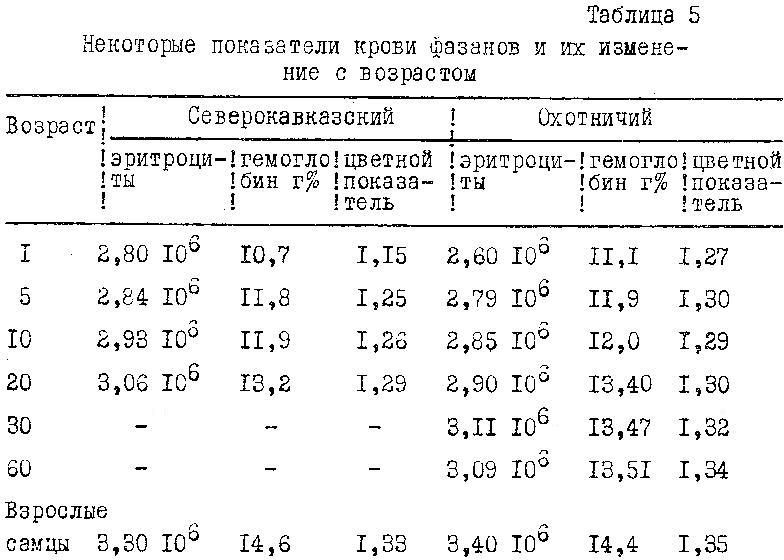
Немаловажное значение приобретает изучение в постэмбриогенезе морфологических и биохимических показателей крови фазанов. Анализ крови охотничьих и северокавказских фазанов, проведенный в те же возрастные периоды, когда определяли массу тела и экстерьер, позволил выявить ряд особенностей в их развитии. С возрастом у фазанов происходило увеличение количества эритроцитов и в соответствии с этим увеличение количества гемоглобина и цветного показателя крови (табл. 5).
Как известно, белки крови играют определенную роль в поддержании онкотического давления, поставляют пластический материал органам и тканям, участвуют в процессах дыхания, иммунитета, обмена углеводов, липидов, витаминов. Динамика уровня белка в крови отражает сдвиги белкового обмена. Результаты исследования содержания общего белка сыворотки крови фазанов представлены в табл. 6.
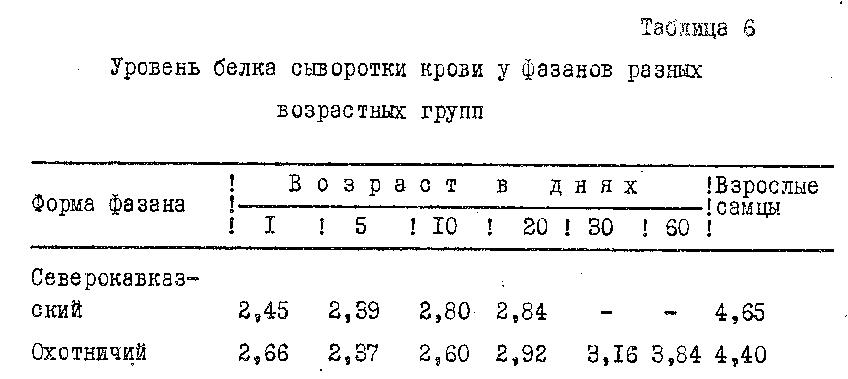
В первые дни постэмбриогенеза у фазанов происходило уменьшение белка в связи с интенсивным его усвоением организмом. Затем, со снижением относительного среднесуточного прироста, происходило увеличение этого показателя. Наибольший показатель белка в крови был у птиц, завершавших свое развитие.
Результаты проведенных исследований крови показали, что на основе детального изучения ее изменения в процессе онтогенеза реальна разработка наиболее простых в исполнении методик прижизненного контроля за развитием фазанов и других видов дичи. Всесторонний контроль за развитием птицы, несомненно, является важнейшей работой на дичефермах. Проводить его по всем показателям (оценка качества суточных фазанят, масса тела, скорость роста, экстерьер, оперяемость молодняка, морфологические и биохимические показатели крови) необходимо в суточном, 10 или 15 (при переводе фазанят из брудера в акклиматизатор), в 80-дневном возрасте и при выпуске в угодья.
И чем раньше начать контроль за развитием птиц, тем быстрее можно выявить воздействие неблагоприятных условий (нарушения режимов кормления, содержания, инкубации и т.д.).
Литература
Второв П.П., Дроздов Н.Н. Определитель птиц фауны СССР. М.; Просвещение, 1980, с. 55-61.
Габузов О.С., Иванова B.C. Перспективы использования фазана в искусственном дичеразведении. - В кн.: Охотничье хозяйство и заповедное дело. М.: Россельхозиздат, 1976, с. 87-95.
Дементьев Г.П. Птицы Советского Союза. М., 1952, т. 4, с. 133-246.
Кузнецов Б.А. Дичеразведение. М.: Лесная промышленность, 1972, с. 178.
Кузьмина М.М.Морфофункциональные особенности задних конечностей куриных. - Тр. ин-та зоологии АН КазССР, 1964, т. 24, с. 152-256.
Кузьмина М.А. Тетеревиные и фазановые СССР. Алма-Ата, 1977, с. 295.
Курскова Т.Н. Периодизация постэмбрионального развития фазана. - Материалы VI Прибалтийской орнитологической конф. Вильнюс, 1966, с. 61-73.
Ларионов В.Ф., Котова О.Д. Биологические закономерности роста цыплят. - Успехи зоотехнических наук, 1936, т. 2, вып. I, с. 63-94.
Шмальгаузен И.И. Проблема пропорцiйного та непропорцiйного росту. 3б. праць Бiол. iн-ту iм. Хв. Омельченко, 1927, №2, с. 77-102.
Шмальгаузен И.И. О факторах роста, определяющих величину тела у птиц. Труды 4-го Всес. съезда зоологов, анатомов и гистологов. Киев-Харьков, 1931, с. 130-131.
Шмальгаузен И.И. Определение основных понятий и методика исследования роста. - В кн.: Рост животных (сб. трудов). М.: Биомедгиз, 1935а, с. 8-60.
Шмальгаузен И.И. Рост и дифференцировка. - В кн.: Рост животных (сб. трудов). М.: Биомедгиз, 1935б, с. 74-84.
Шмальгаузен И.И. Рост и общие размеры тела в связи с их биологическим значением. - В кн.: Рост животных (сб. трудов). М.: Биомедгиз, 1935в, с. 61-73.
Nady Е.A. A facan es a fogoly intenziv tenyesztese. - Budapest, 1971, 224 p.
Heinroth O., Heinroth M. Die Vogel Mitteleuropas. Berlin, 1924-1932, Bd.1-4, 312 s.