




Корж А.П. К особенностям периодизации постэмбрионального развития птенцов охотничьего фазана в условиях юга Украины. Сообщение 2. // Вестник зоологии № 35 (2). С. 93-97. 2001 г.
К ОСОБЕННОСТЯМ ПЕРИОДИЗАЦИИ ПОСТЭМБРИОНАЛЬНОГО РАЗВИТИЯ ПТЕНЦОВ ОХОТНИЧЬЕГО ФАЗАНА В УСЛОВИЯХ ЮГА УКРАИНЫ
СООБЩЕНИЕ 2
Зависимость постэмбрионального развития фазана от условий выращивания затрудняет построение периодизации, которую можно было бы применять на практике. Нами выделены 3 периода постэмбриогенеза: до 30-дневного возраста — период развития; до 90-дневного возраста — период роста и после 90-дневного возраста — период взросления. В зависимости от условий и генотипа выращиваемой птицы, с учетом географической зоны, появление отдельных признаков колеблется в пределах нескольких недель, поэтому сроки выделенных периодов нельзя считать жестко детерминированными. При оптимальных условиях выращивания первый период может заканчиваться к 21-му дню, а второй — к 60-му дню выращивания.
Ключевые слова: охотничий фазан, рост, развитие, постэмбриогенез, периодизация.
Some Peculiarities of Periodization of Hunting Pheasant Postembriogenesis in Southern Ukraine. Communication 2. Korzh A. P. — A strong dependence of a postembrional development of the hunter pheasant on conditions of breading makes it difficult a periodisation, which could be used in practice. We have defined three periods of the postembriogenesis: up to 30 days age, the period of development; up to 90 days age, the period of growth; and after 90th day, the period of adulting. Depending on conditions and a genotype of breading bird and a geographical zone as well the appearance of some characters range around the limits of some weeks. Therefore, the terms of defined periods cannot be considered as exactly determined. Under optimal conditions of breading a first period can finish by the 21st day and the second — by the 60th. Key words: hunter pheasant, growth, development, postembriogenesis, periodisation.
Введение
Единая, общепризнанная периодизация постэмбрионального развития фазана пока отсутствует, а разные авторы подходят к решению этой проблемы неодинаково. Периодизация Т.Н. Курсковой (1965) основывается в первую очередь на характеристике общего весового роста. Работы Б.А. Кузнецова (1972) и С.Л. Курочкина (1985) учитывают некоторые экологические моменты в жизни птенцов. К. Лаубе (Laube, 1979) делает значительный акцент на особенности развития перьевого покрова фазанят, а В.С. Иванова и Н.Н. Трошкина (1983, 1986) без разработки периодизации как таковой предлагают контрольные величины показателей для того или иного возраста.
Серия специальных экспериментов по изучению влияния разных условий содержания и кормления на рост молодняка фазана (Корж, 1998) позволила сделать ряд обобщений, касающихся влияния различных факторов на развитие и физиологическое состояние птицы. Основываясь на результатах этих исследований, а также на анализе литературных данных, можно предложить упрощенный, но удобный в практическом использовании вариант периодизации постэмбриогенеза фазана.
Материал и методы
Исследования проводились в 1989-1994 гг. на базе Отрадновского охотничьего хозяйства Запорожской обл. и фазанария «Холодная гора» Республики Крым. Исследовалось влияние различных условий содержания и кормления на рост и развитие фазанят (Корж, 1998). Математическая обработка полученных результатов проводилась по общепринятым методикам (Лакин, 1990).
Результаты и обсуждение
Разногласия в выделении периодов постэмбриогенеза существуют не только по поводу охотничьего фазана, но и для других организмов. Г.А. Шмидт (1968) считает, что при построении периодизации онтогенеза любого организма необходимо опираться на типы его экологических взаимоотношений с окружающей средой. На наш взгляд, этот подход возможен только для выделения наиболее крупных этапов онтогенеза. По крайней мере, для куриных характерно вылупление птенцов с уже полностью сформированными органами и системами органов, после чего происходит только их становление (Фисинин и др., 1990). Более того, Л.П. Познанин (1979) приходит к выводу, что для птиц в постэмбриогенезе выделение четких периодов практически невозможно. Для фазана это осложняется еще его гибридогенным происхождением, что обусловливает значительный полиморфизм (Искусственное разведение..., 1987; Luders, 1989), и сильной зависимостью ростовых процессов от условий выращивания (Солоха, 1987, 1989; Lange, Rizzi, 1988; Woodard et al., 1977).
Возможно, что именно эти факты обусловливают противоречия существующих периодизаций (сроки смены перьевого покрова, появление полового диморфизма, становление пропорций тела взрослого организма и т.д.). При этом, чем подробней периодизация постэмбрионального развития, тем больше отклонений от нее наблюдается на практике. С.С. Шварц (1973), например, утверждает, что в любом биологическом единстве изменения свойств одного из его составляющих меняет поведение системы в целом. Организм, который также может рассматриваться как биологическая система, при изменении каких-либо факторов не может не изменить хода своего онтогенеза. В Сообщении 1 (Корж, 1998) приводятся данные об ускорении или замедлении роста фазанят в зависимости от условий их выращивания.
Несмотря на значительные отклонения в развитии отдельных признаков в зависимости от условий выращивания, была установлена устойчивость общего хода постэмбриогенеза фазана. Корреляционный анализ показал, что общая закономерность развития птенцов сохраняется практически без изменений при их выращивании в разных условиях — коэффициент корреляции между развитием линейных характеристик в разных условиях не опускался ниже 0,97, что значимо при Р<0,001. Поэтому можно говорить о канализированности процессов развития птенцов фазана.
Было установлено, что развитие молодняка по отдельным показателям подчиняется параболической закономерности. При этом динамика набора массы тела имеет типичную S-образную форму (рис. 1), что согласуется с данными других авторов (Мина, Клевезаль, 1976; Солоха, 1987). Линейные параметры также наиболее достоверно описываются параболической закономерностью (коэффициенты детерминации для параболической закономерности были наибольшими и составляли 94,6-99,5%). Эти данные совпадают с выводом И.И. Шмальгаузена (1984) о том, что экспоненциальная и логистическая кривые не отражают закономерностей параболического роста организмов, который является результатом суммирования параболических кривых роста органов и тканей. В то же время, М.В. Мина и Т.А. Клевезаль (1976) отмечают, что само по себе изменение соотношения массы и линейных размеров может в ряде случаев дать основание для важных экологических и физиологических обобщений.
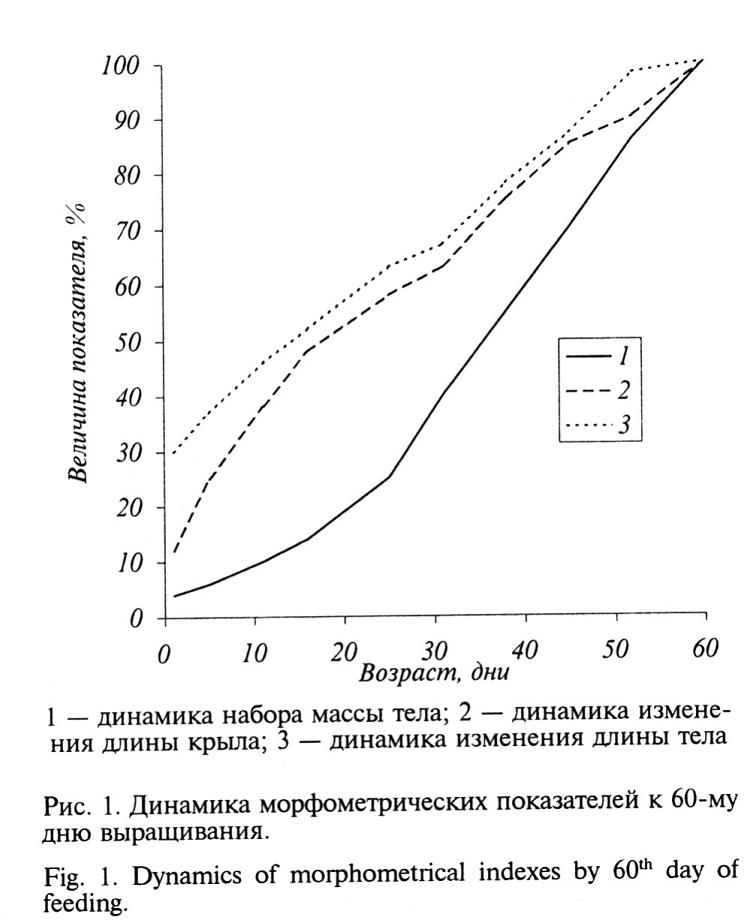
Исходя из этого, наиболее целесообразным представляется построение периодизации постэмбриогенеза фазана на основе общебиологических закономерностей. Так, в онтогенезе некоторых птиц довольно четко прослеживаются 3 основных этапа роста, отличающиеся особенностями ростовых процессов. Первый период — максимального роста, второй — меньшей скорости роста, и третий — слабое увеличение или потеря массы тела. Последний период рассматривается как время подготовки к полету птиц, сразу после выхода из гнезда становящихся на крыло (Мина, Клевезаль, 1973).
Несмотря на то, что фазан относится к выводковым (матуронантным) птицам, его ростовые процессы также отличаются определенной периодичностью. Исходя из этого, наиболее рациональным, на наш взгляд, является выделение 3 основных периодов постэмбриогенеза фазана без их дальнейшей детализации.
Первый период, продолжающийся с 1-го по 30-й день, характеризуется наиболее интенсивными темпами роста птенцов по всем показателям — особенно в первые 10-15 дней (рис. 2). Наиболее активно идет развитие линейных характеристик птенцов в сравнении с набором массы тела. В оптимальных условиях выращивания к концу периода у птенцов завершается формирование ювенильного оперения и системы терморегуляции, в связи с чем отпадает необходимость в дальнейшем обогреве. В зависимости от условий выращивания масса тела птенцов составляет 140-200 г, что не превышает 10-15% массы тела взрослой птицы.
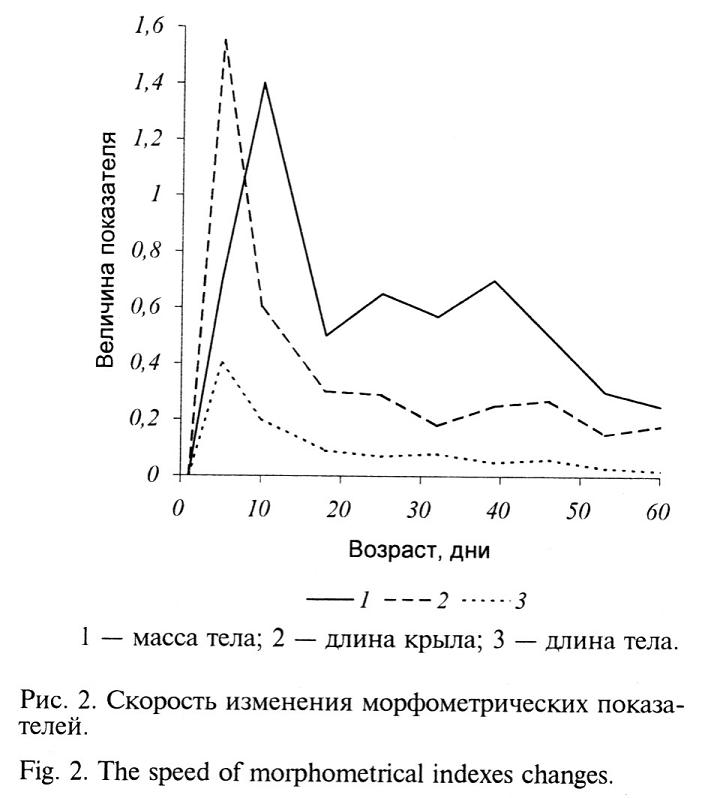
Поскольку формирование гомойотермии идет по алломорфному пути, одновременно с ростом мускулатуры и оперения, необходимых для подготовки полетной активности (Давидов, Кекспайк, 1992), увеличение массы тела в этом периоде следует рассматривать как результат развития организма в целом. Содержание фазанят в неблагоприятных условиях приводит к явному отставанию в развитии, поскольку в этом периоде они наиболее восприимчивы к воздействию неблагоприятных факторов. Чем оптимальней условия выращивания, тем более стабильна и равномерна динамика отдельных показателей роста и развития птенцов. Но даже в этом случае коэффициент вариации редко опускается ниже 10%.
Во втором периоде, длящемся с 31-го по 90-й день, развитие оперения и скелета постепенно замедляется, но наблюдается интенсивный набор массы тела (к 90-дневному возрасту отдельные экземпляры могут достигать килограмма и более). Развитие во втором периоде более неравномерно, чем в первом, что отмечают и другие исследователи (Курочкин, 1985; Солоха, 1987, 1989). В норме, к концу этого периода у птицы полностью завершается формирование дифинитивного оперения, масса тела сопоставима с таковой у взрослой птицы, ростове процессы практически завершаются. При неблагоприятных условиях выращивания наблюдается значительное отставание в наборе массы тела и формировании дифинитивного оперения.
В третьем периоде, начинающемся после 90-го дня, отмечается постепенное торможение ростовых процессов и завершается половое созревание птицы. В этом периоде довольно часто наблюдается продолжение роста отставших в развитии птенцов. При неблагоприятных условиях выращивания животные не достигают максимально возможных размеров, а в дальнейшем снижается их продуктивность. Это согласуется с литературными данными о том, что если стресс был тяжелым и его воздействие пришлось на ранние периоды жизни, то наверстывающий рост может оказаться неполным (Мина, Клевезаль, 1976; Теннер,1979). Возможно, именно этот факт обусловливает постепенное вырождение той или иной популяции.
Поскольку становление терморегуляции и развитие первичного перьевого покрова заканчивается лишь к концу первого периода, подразделять его на более мелкие этапы нецелесообразно. Выделение первых 2-3 дней постнатального развития в отдельный период (Кузнецов, 1972; Курочкин, 1985; Рахманов, Бессарабов, 1991), по нашему мнению, лишено оснований. К моменту вылупления основные физиологические системы цыпленка сформированы, а их становление происходит в течение 1-2 недель постнатальной жизни, о чем свидетельствует повышенная смертность в этот период (Искусственное разведение..., 1987; Фисинин и др., 1990). Одновременно с этим происходит и становление терморегуляции организма (Давидов, Кекспайк, 1992).
Л.П. Познанин (1979) указывает, что следует разграничивать такие понятия, как стадия и этап развития; при этом каждый этап ограничивается и отделяется от других определенными стадиями развития. Определенную сложность периодизации вызывает и большая зависимость ростовых процессов охотничьего фазана от условий выращивания. Подобной точки зрения придерживаются и другие авторы — выделять периоды роста следует осторожно, поскольку торможение ростовых процессов может быть вызвано исключительно внешними причинами, а недостаток каких-либо условий для нормального развития организма может быть компенсирован в дальнейшем наверстывающим ростом (Мина, Клевезаль, 1976; Теннер, 1979).
Сроки начала и завершения предложенных нами периодов не являются строго детерминированными и могут сдвигаться в зависимости от условий выращивания. При условии доброкачественности кормов, высоком уровне протеина в них, а также соблюдении нормативов плотности посадки птенцов, первый период может заканчиваться к 21-му дню, а второй — к 60-му дню выращивания.
Выводы
Несмотря на существенное влияние условий содержания и кормления на общий ход онтогенеза птенцов фазана, процессы его развития могут считаться канализированными.
Наиболее целесообразным является выделение 3 крупных периодов постэмбрионального развития птенцов фазана.
При благоприятных условиях выращивания, продолжительность первого и второго периодов может существенно сокращаться. Отставание в развитии может частично или полностью компенсироваться последующим наверстывающим ростом.
Список литературы
Давидов А. Ф., Кекспайк Ю.Э. Онтогенез, терморегуляция у птиц. — С.-Пб. : Наука, 1992. 176 с.
Иванова В.С., Трошкина Н.Н. Контроль за развитием обыкновенного фазана // Искусственное разведение фазанов. М., 1983. С. 128-145.
Иванова В.С., Трошкина Н.Н. Методические рекомендации по контролю за развитием молодняка птиц, выращиваемых в искусственных условиях. М., 1986. 10 с.
Искусственное разведение фазанов / Под ред. О.С. Габузова. М., 1987. 140 с.
Корж А.П. К особенностям периодизации постэмбрионального развития птенцов охотничьего фазана в условиях юга Украины. Сообщение 1. Влияние различных факторов на постэмбриогенез охотничьего фазана // Вестн. зоологии. 1998. 32, № 5-6. С.115-118.
Кузнецов Б.А. Дичеразведение. М.: Лесная промышленность, 1972. 180 с.
Курочкин С.Л. Особенности постэмбрионального развития обыкновенного фазана в Северо-Западном Причерноморье // Искусственное дичеразведение. М., 1985. С. 85-101.
Курскова Т.Н. Рост и развитие фазана и применение антибиотиков при его разведении : Автореф. дисс. ... канд. биол. наук. Минск, 1965. 18 с.
Мина М.В., Клевезаль Г.А. Рост животных. М.: Наука, 1976. 291 с.
Познанин Л. П. Эколого-морфологический анализ онтогенеза птенцовых птиц. М.: Наука, 1979. 293 с.
Рахманов А.И., Бессарабов Б.Ф. Фазановые: содержание и разведение. — М. : Агропромиздат, 1991. 174 с.
Солоха А.В. Искусственное разведение хивинского фазана // Разведение ценных и редких видов животных. М., 1987. С. 14-30.
Солоха А.В. Особенности биологии некоторых среднеазиатских подвидов фазана при искусственном разведении // Зоокультура ценных и редких видов птиц и зверей. М., 1989. С. 62-82.
Тэннер Дж. Рост и конституция человека // Биология человека. М.: Мир, 1979. С. 366-472.
Фисинин В.И., Журавлев И.В., Айдинян Т.Г. Эмбриональное развитие птицы. М. : Агропромиздат, 1990. 240 с.
Шварц С.С. Эволюция биосферы // Проблемы биогеографии. М.: Наука, 1973. С. 213-228.
Шмалъгаузен И.И. Рост и дифференцировка. Киев : Наук. думка, 1984. 2. 166 с.
Шмидт Г.А. Типы эмбриогенеза и их приспособительное значение. М.: Наука, 1968. 230 с.
Laube К. Alters-, Rassen- und Gaschlechtsmercmale beim Fasanenkiiken und Jungfasanen // Dis. ... — Giessen, 1979. - 81 Р.
Lange E., Rizzi S. Alimentarione Tradicionale e racionale del fagiano // Rivista di zootechnica. 1966. 39, N 11. P. 587-592.
Luden H. Was ist bei der Zucht, Haltung und Futterung von fasanen zu beachten? // DGS. 1989. N 13. Р. 547-551.
Woodard A.E., Vohra P., Snyder R.L. Effect of Protein Levels in the Diet on the Growih of Pheasants from // Poultry sciense. 1977. 56, N 5. P. 1492-1500.