




Павлющик Т.Е., Малютина Н.В. Некоторые морфологические характеристики глухарей, выращенных в искусственных условиях // Разведение ценных и редких видов животных. Сборник научных трудов ЦНИЛ Главохоты РСФСР. Москва, 1987. С. 55-61.
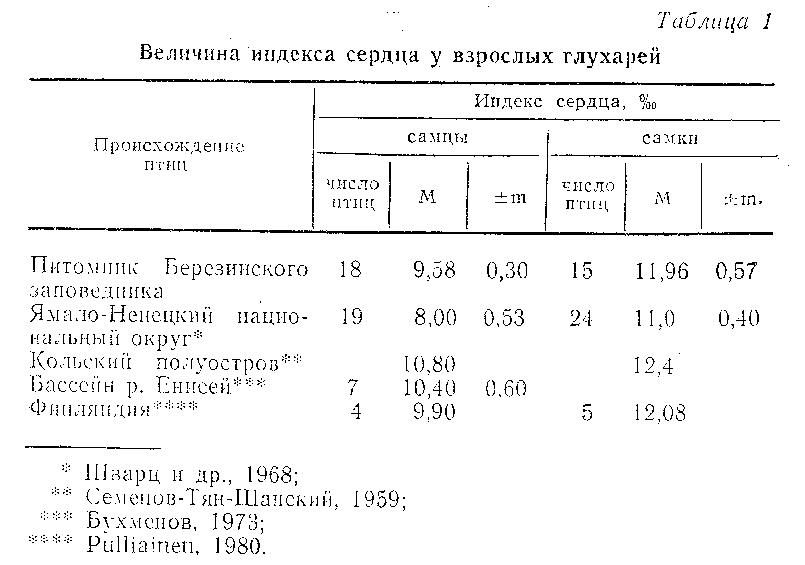
В работе рассматриваются такие важные характеристики, отражающие состояние глухарей, выращенных и содержавшихся в неволе, как индексы сердца, мускульного желудка и различных отделов кишечника. Эти показатели определялись по общепринятой методике (Шварц и др., 1968) для птиц, погибших в питомнике Березинского заповедника в 1972-1984 гг. Пол и возраст глухарей был известен.
Сердце. Абсолютная и относительная масса сердечной мышцы характеризует уровень двигательной активности животного. Хотя гипертрофия миокарда не всегда является единственным ответом сердечно-сосудистой системы на повышение активности, основная закономерность здесь такова: чем большая работа совершается сердцем в единицу времени, тем резче выражена его гипертрофия (Шварц и др., 1968; Дофинский, 1981). Таким образом, изменение абсолютной и относительной массы сердца у животных, содержащихся в неволе, в сравнении с дикими может служить показателем их двигательной активности и свидетельствовать об адекватности условий содержания.
Глухари в питомнике Березинского заповедника содержались в ограниченных по площади вольерах и клетках (соответственно 20-35 и 9 м2), причем максимальная продолжительность жизни в неволе составила 11 лет. Сопоставление индекса сердца глухарей из питомника и диких птиц (табл. 1) свидетельствует о том, что изменчивость этого показателя у птиц из питомника не выходит за пределы нормальной видовой изменчивости данного органа. Следовательно, при содержании в неволе даже при ограничении площади клетки до 9 м2 двигательная активность глухарей сопоставима с таковой в природных условиях. Это вполне объясняется тем, что физиологическая потребность данного вида в движении невелика: глухарь в природе ведет довольно малоподвижный образ жизни,проводя в полете, особенно в зимнее время, всего несколько минут в сутки (Семенов-Тян-Шанский 1959; Потапов, 1982). В семействе тетеревиных глухарь имеет один из самых низких сердечных индексов, уступая только рябчику (Шварц и др., 1968).
Мускульный желудок. Среди выращенных в питомнике глухарей максимальная масса желудка обнаружена у самцов в возрасте 6-11 лет: 110,4±10,8 г. У самок старше одного года возрастные изменения массы желудка отсутствуют. Сравнение относительной и абсолютной массы мускульного желудка глухарей из питомника и диких птиц (табл. 2) показало, что эти величины у птиц из питомника всегда ниже, чем в природе. Только у активно токующих самцов бассейна р. Енисея индекс желудка был ниже, чем у глухарей в неволе (Бухменов, 1973).
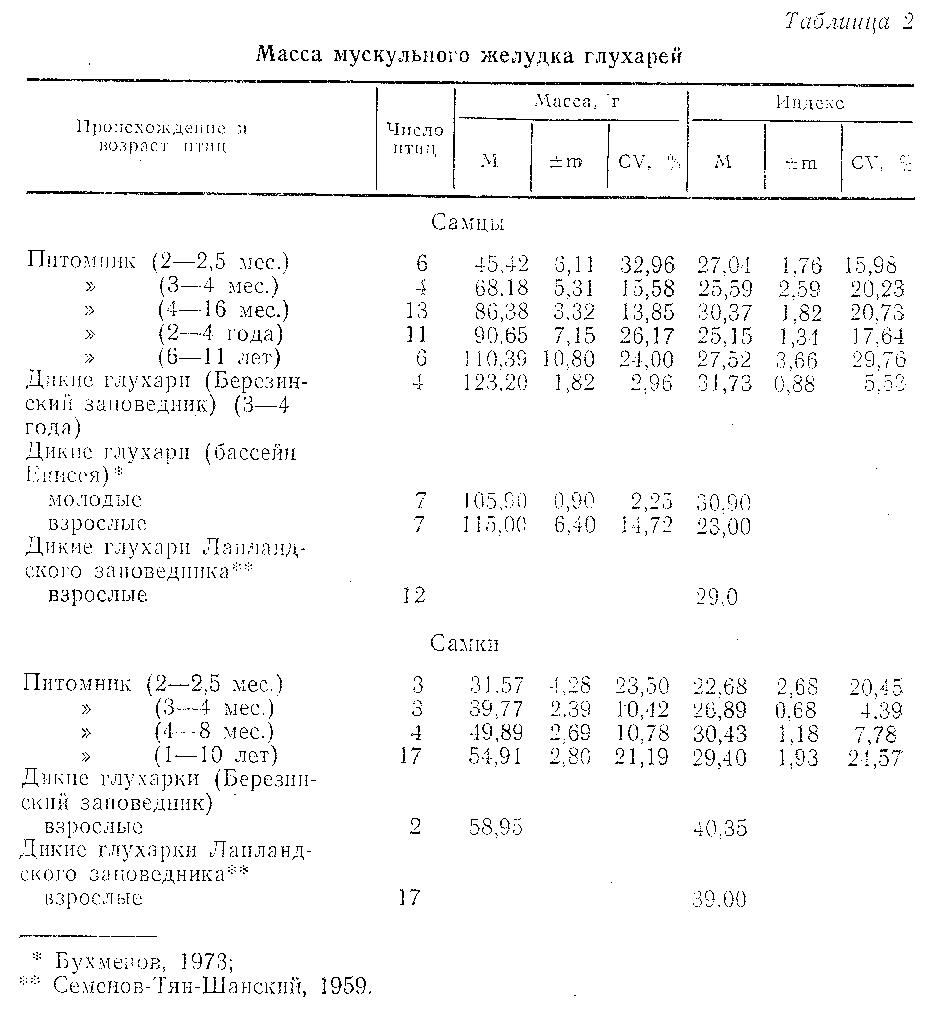
Если принять за 100 % массу мускульного желудка у диких глухарей Березинского заповедника, то уменьшение ее у птиц из питомника составило 10,4 % для самцов и 8,5 % для самок, что значительно ниже сезонных вариаций массы данного органа у тетеревиных птиц (Семенов-Тян-Шанский, 1959). В наших опытах глухари в неволе потребляли искусственные корма со средним содержанием сырой клетчатки 15 % от сухого вещества, что привело к сравнительно небольшому, по стабильному уменьшению массы мускульного желудка в сравнении с дикими птицами. Поскольку это уменьшение не выходит за пределы сезонных изменений массы мускульного желудка диких птиц, связанных с сезонными изменениями их рационов, можно предположить, что оно не будет существенным препятствием для перехода глухарей, выращенных в неволе, на естественные корма в случае их выпуска в природу.
Кишечник. Суммарная длина тонких и прямой кишок у птиц из питомника представлена в табл. 3, слепого отдела кишечника - в табл. 4. Комментируя эти таблицы, необходимо отметить, что у глухарей, получавших искусственные корма, наибольшие изменения произошли именно в слепом отделе кишечника. Как известно, длина слепых кишок - признак адаптивный, зависящий от рациона и от продолжительности зимнего периода, т.е. от длительности употребления тетеревиными птицами грубых кормов (Потапов, 1974). Длина слепых кишок варьирует у разных популяций внутри вида и у данной популяции в разные сезоны, причем северные популяции обычно имеют большую длину слепого кишечника, чем обитающие в южных широтах (Потапов, 1974; Jasaway, 1976).
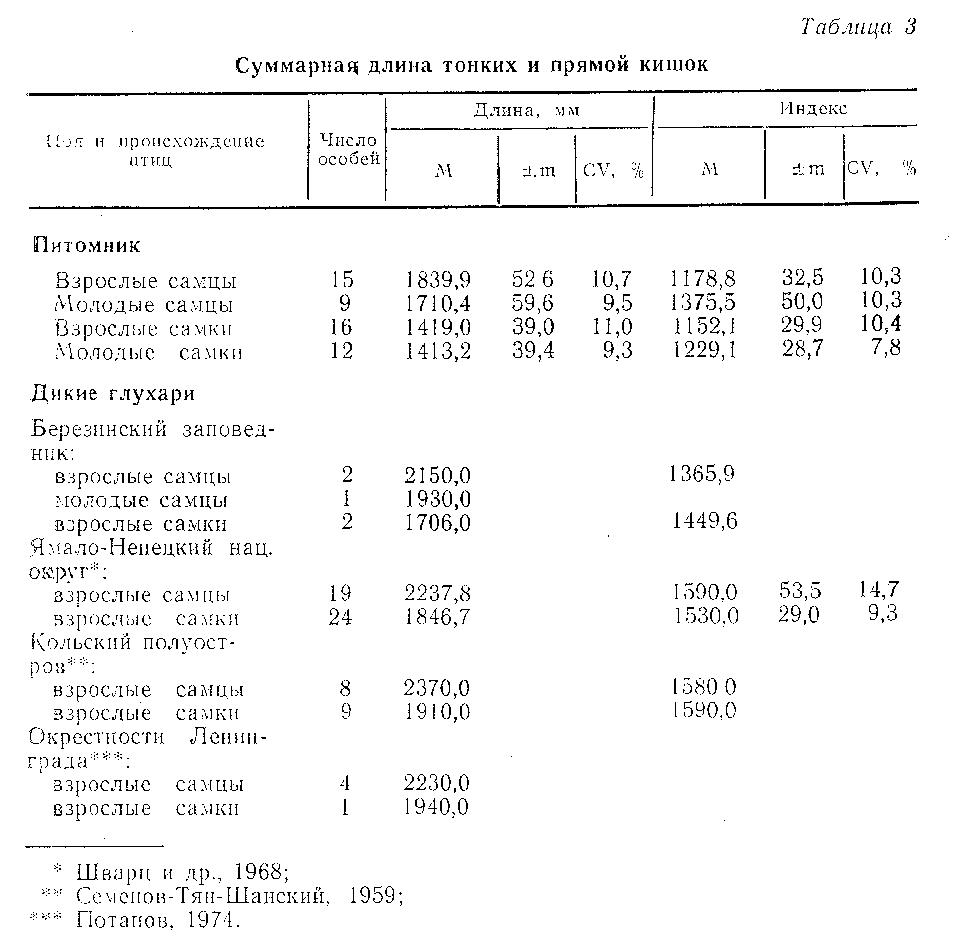
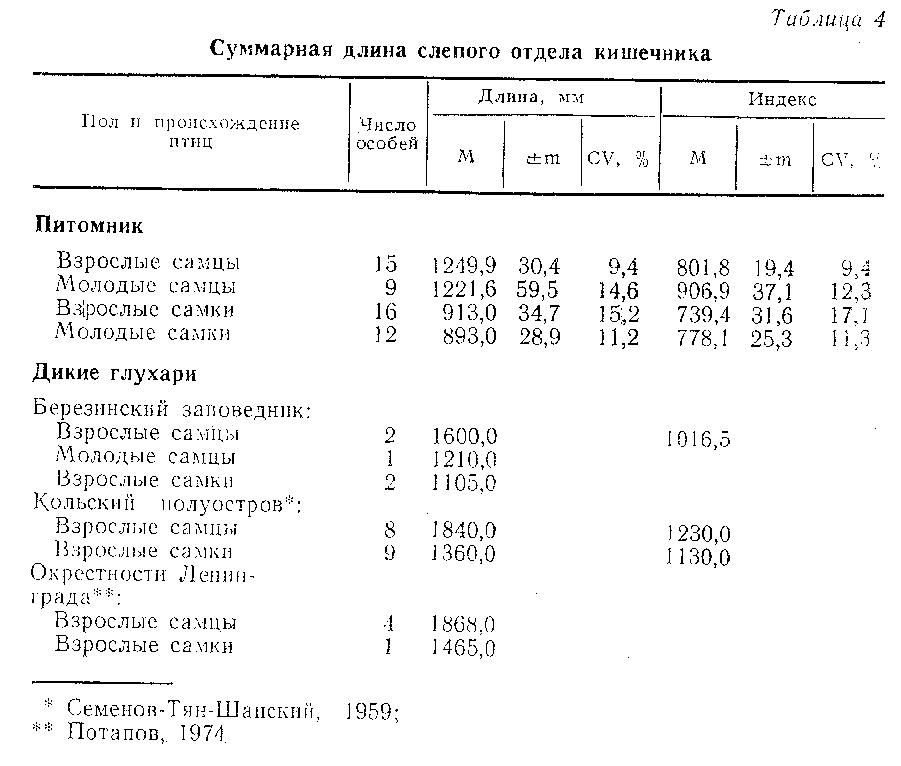
Дикие глухари Березинского заповедника имеют меньшую длину всех отделов кишечника в сравнении с птицами, обитающими севернее. Однако даже в сравнении с птицами местной популяции уменьшение общей длины кишечника у глухарей из питомника составляет 16,8 %, а слепого отдела - 17,4 % для самок и соответственно 14,4 и 21,9 % для самцов. Низкая вариабельность как абсолютных, так и относительных показателей длины различных отделов кишечника указывает на их стабильность и косвенным образом свидетельствует о сохранениями значимости процессов пищеварения в слепом отделе кишечникадля глухарей, потребляющих в неволе искусственные рационы. Для сравнения укажем, что коэффициент вариации слепого отдела кишечникау диких глухарей составляет 11 % (Добринский, 1981), следовательно, вариабельность этого показателя у птиц из питомника сопоставима с нормальной для вида. Нормальные сезонные вариации в длине слепых кишок составляют примерно 11,6 % (Jasaway, 1976), следовательно, полученные в наших экспериментах результаты выходят за пределы этих вариаций.
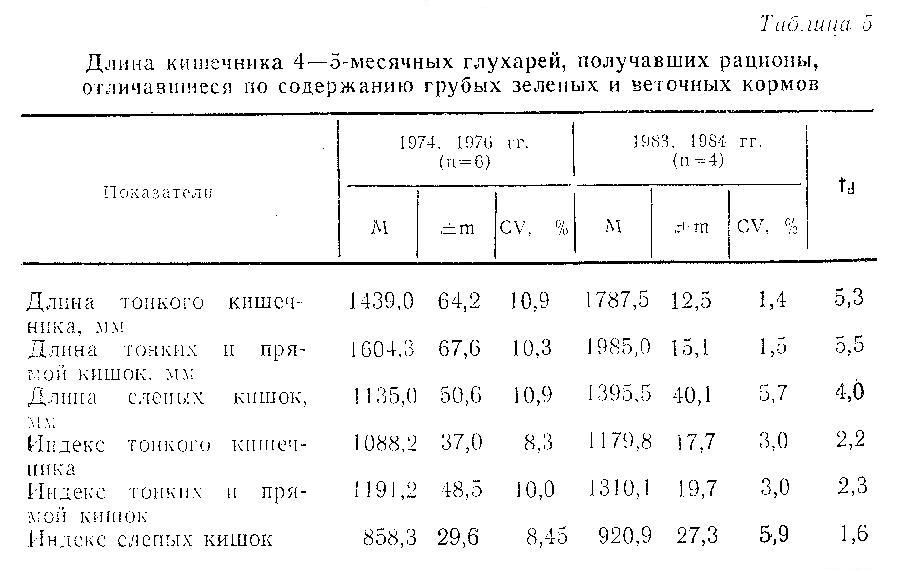
Одним из важнейших факторов, вызывающих сокращение длины кишечника у тетеревиных птиц, разводимых в неволе, является низкое в сравнении с естественными кормами содержание клетчатки в рационах. Так, в экспериментах на шотландской куропатке, получавшей искусственные рационы с 11-12 % сырой клетчатки, сокращение длины слепого отдела кишечника составило 48 %, а тонких и прямых кишок – 28 % (Moss, 1972), что почти в 2 раза превосходит полученные нами результаты при содержании в рационе примерно 15 % сырой клетчатки. Мы предположили, что, изменяя рационы содержащихся в неволе птиц в сторону увеличения содержания грубых зеленых и веточных кормов, можно до некоторой степени скорректировать негативное влияние искусственных рационов на длину кишечника. С этой целью мы сопоставили длину кишечника у молодых глухарей, выращенных в питомнике и получавших рационы, отличающиеся по содержанию сырой клетчатки (табл. 5). Птицы, выращенные в 1983-1984 гг., получали с 40-дневного возраста грубые зеленые корма: вико-овсяную смесь и желтый люпин. Хвою сосны они начали потреблять в 70-дневном возрасте. Глухари, выращенные в 1974 и 1976 гг., получали в течение всего периода выращивания мягкую огородную зелень. Хвоя сосны была включена в рацион с 90-дневного возраста. У глухарей 1983-1984 гг. отмечено статистически достоверное увеличение абсолютных размеров кишечника в сравнении с группой, рацион которой был ограничен по сырой клетчатке. Относительные размеры кишечника также оказались выше у птиц 1983-1984 гг., однако статистически достоверные различия получены только для индекса суммарной длины тонкого и прямого отделов кишечника, что, возможно, связано с малой величиной выборки. Проведенное сравнение указывает на возможный путь совершенствования рационов глухарей при разведении в неволе с целью приближения морфологических характеристик пищеварительного тракта к норме, свойственной диким птицам.
Выводы
1. Величина индекса сердца у глухарей, содержащихся в
неволе, не выходит за пределы нормальной видовой изменчивости данного показателя, что свидетельствует об адекватности условий содержания глухарей их физиологической потребности в движении.
2. Уменьшение массы мускульного желудка у глухарей, потребляющих искусственные корма, содержащие около 15 % сырой клетчатки, составляет 10,4 % для самцов и 8,5 % для самок в сравнении с дикими птицами местной популяции, что ниже сезонных вариаций массы данного органа.
3. Уменьшение длины кишечника у глухарей, содержащихся в неволе, может быть скорректировано путем увеличения содержания сырой клетчатки в рационе за счет грубых зеленых и веточных кормов.
ЛИТЕРАТУРА
Бухменов В.П. О соотношении половой активности глухарей с их морфо-физиологическими особенностями // Экология, 1973. № 3. С. 94.
Добринский Л.Н. Динамика морфо-физиологических особенностей птиц. М.: Наука, 1981.
Потапов Р.Л. Адаптации семейства Tеtraonidae к зимнему сезону // Труды ЗИН АН СССР. Л., 1974. Т. 55. С. 207-251.
Потапов Р.Л. Биоэнергетика тетеревиных птиц в зимний период // Труды ЗИН АН СССР. Л., 1982. Т. 113. С. 57-67.
Семенов-Тян-Шанский О. И. Экология тетеревиных птиц. М., 1959.
Шварц С.С., Смирнов В.С, Добринский Л.Н. Метод морфо-физиоло-гических индикаторов в экологии наземных позвоночных // Труды Института экологии растений и животных Уральского филиала АН СССР. Свердловск. 1968. Вып. 58. С. 1-387.
Jasaway W. Seasonal variations in diet, volatile fatty acids production and size of the cecum of Rock ptarmigan // Соmр. Biochem. Physiol., 1976, 53A. P. 109-114.
Moss R. Effects of captivity on gut length in Red Grouse // Wildlife Management, 1972. V. 36. № 1. p. 99-104.
Pulliainen E. Heart-body weight ratio in the Willow grouse and Rock Ptarmigan in Finnish Lapland // Orris Fennica, 1980. V. 57. P. 88-90.