




Трошкина Н.Н., Иванова В.С. Сравнительное изучение постэмбрионального роста двух групп канадских казарок // Дичеразведение в охотничьем хозяйстве. Сборник научных трудов ЦНИЛ Главохоты РСФСР. Москва, 1985. С. 69-81.
Идея акклиматизации большой канадской казарки в СССР принадлежит В.Ф. Гаврину. В 1970 г. на Межведомственном совещании по гусям и казаркам (г. Тарту) он выступил с идеей интродукции ее в нашей стране. В 1973 г. на Втором Всесоюзном совещании по акклиматизации и реакклиматизации охотничьих животных в СССР это предложение было занесено в решение совещания.
Перед отделом дичеразведения ЦНИЛ Главохоты РСФСР была поставлена задача разработать технологию искусственного разведения канадских казарок на фермах.
Для первых поисковых экспериментов яйца канадской казарки были получены из Московского и Каунасского зоопарков. В 1976 и 1978 гг. птиц приобрели за рубежом. Данное поголовье канадских казарок, находившееся в дичепитомнике ЦНИЛ, было использовано при разработке технологии массового разведения этого вида с использованием промышленных приемов содержания, кормления и разведения. Критерием, позволяющим оценивать применяемые методы и способы содержания и кормления родительского поголовья, является качество молодняка, которое характеризуется ростом массы тела и гармоничностью сложения. Для контроля правильности выбранных нами методов выращивания молодняка канадских казарок в дичепитомнике ЦНИЛ проводили сравнение их развития с развитием молодняка, выведенного из яиц канадских казарок, поступивших из Слимбриджа (Англия). В 1980 г. ЦНИЛ получила из Слимбриджа 18 яиц канадской казарки. Яйца были уже насиженными (зародыши находились на последней стадии развития). Из яиц было выведено и выращено в дичепитомнике ЦНИЛ 8 птиц. Выращивание молодняка двух опытных групп канадских казарок - 25 птенцов, вылупившихся из яиц, полученных в дичепитомнике ЦНИЛ (I группа), и 8 птенцов из яиц, полученных из Англии (II группа) - происходило в идентичных условиях по методикам, разработанным в отделе дичеразведения. В основу исследований было положено изучение массы тела и экстерьерных показателей в постэмбриогенезе. Для более полного отражения изменений экстерьера птиц проанализировано 15 морфологических показателей в разные возрастные периоды (1, 5, 10, 20, 30, 60, 90, 360 дней). Молодняк, после того как он обсыхал, индивидуально метили крылометками. Выведенные из полученных в ЦНИЛ яиц птенцы канадской казарки (I группа) имели массу от 115 до 123 г, что составляло 65 % массы яиц до инкубации, птенцы же, полученные из яиц, присланных из Англии (II группа), - от 99 до 120 г, что составляло 73,9 % от массы яиц до инкубации. Исследования проводили с учетом пола каждой особи. Рост молодняка канадской казарки двух групп проиллюстрирован на рис. 1 и 2. Как видно, нарастание массы тела у канадских казарок продолжалось до 360 дней. Кривые роста самцов до 60 дней не отличались, а далее до 90 дней самцы II группы опережали самцов I группы, однако после 90 дней темп их роста сравнялся; у самок, начиная с 20-дневного возраста, более интенсивный рост наблюдался у птиц II группы, однако, эти различия у годовалых птиц не были достоверны.
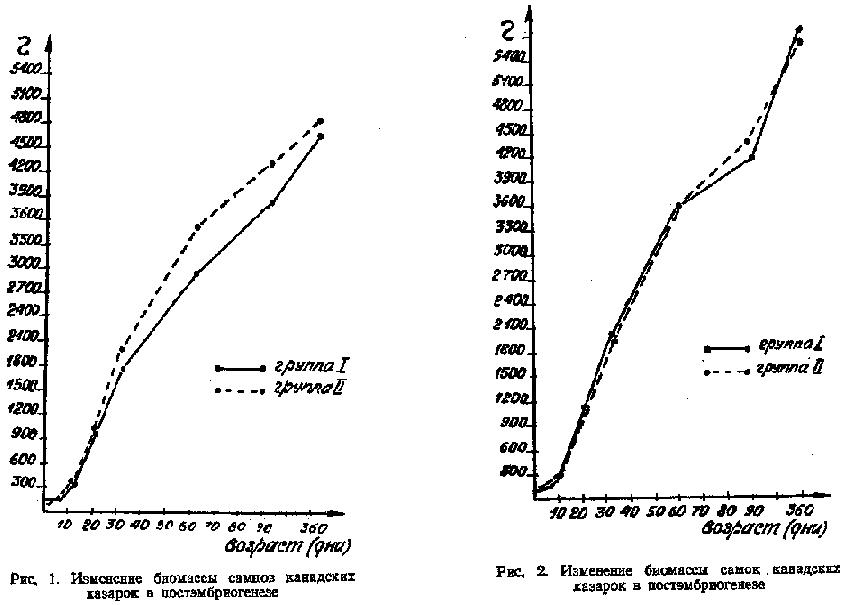
При изучении полового диморфизма по массе тела было установлено, что у 30-дневных самцов канадских казарок I группы масса достоверно отличалась от массы самок, которая составляла 88 % от массы самцов, а во II группе достоверные отличия выявлены лишь в 60-дневном возрасте, когда масса самок составляла 95 % от массы самцов.
У обеих групп птиц было проведено изучение роста в постэмбриогенезе таких частей тела, как голова, туловище и конечности. В табл. 1 приведена динамика постэмбрионального роста головы канадской казарки по трем основным направлениям: длине, ширине и высоте. Сравнение всех трех показателей у суточных птенцов и у годовалых птиц показало, что у молодняка голова наиболее быстро растет в ширину (48-56 % ширины головы взрослых птиц), несколько хуже она росла в высоту (45-49 %) и менее всего в длину (38-40 %). Наиболее интенсивный рост головы происходит в первый месяц постэмбриогенеза, к его концу размеры головы птенцов достигали уже 70-93 % размеров головы у взрослых птиц, причем соотношение показателей почти не изменилось, то есть голова больше выросла в ширину (81-93 %), несколько меньше в высоту (71-82 %) и менее всего в длину (71-76 %). У двухмесячных птиц размеры ее достигли уже 90-99 % размеров взрослых птиц, причем ширина головы выросла до 95-99 %, высота - до 91-93 % и длина - до 90-96 %. У трехмесячных птиц рост головы почти завершился, ее размеры составляли уже 95-100 % размера годовалых птиц. Длина головы суточных птенцов II группы превышала таковую I группы на 1-2 мм, и эти незначительные отличия прослеживались до 90 дней. Однако, в возрасте 360 дней, в отличие от суточных птиц, длина головы у казарок I группы была на 2 мм больше, чем у птиц II группы. Это различие статистически недостоверно. Если у суточных птенцов I и II групп отличий в ширине и высоте головы не обнаружено, то в результате разного темпа роста месячные птенцы I группы (как самцы, так и самки) имели ширину на 5 мм больше, а высоту - наоборот, на 3-4 мм меньше, чем птенцы II группы. На последующих этапах постэмбриогенеза это различие нивелируется за счет более интенсивного роста головы в ширину у птиц II группы и в высоту у птиц I группы. Годовалые птенцы I группы имели ширину и высоту головы на 1-2 мм больше, чем II группы, однако это отличие статистически недостоверно.
Рост клюва канадских казарок в длину и ширину дан в табл. 2. Длина клюва представлена тремя промерами: длиной от основания, от ноздри и от угла рта. У только что вылупившегося птенца длина клюва составляла от 27 до 32 %, а ширина - от 35 до 40 % соответствующих показателей взрослых птиц. За первые 30 дней отмечается наибольший (за весь период постэмбриогенеза) рост клюва; его длина у месячных птиц составила 65-76 %, а ширина 74-83 % от размеров клюва взрослых птиц. В последующие 30 дней рост клюва в длину происходил интенсивнее, чем в ширину, и у 2-месячных птиц его длина достигала 86-93 %. К 3 месяцам рост клюва как в длину, так и в ширину почти завершился.
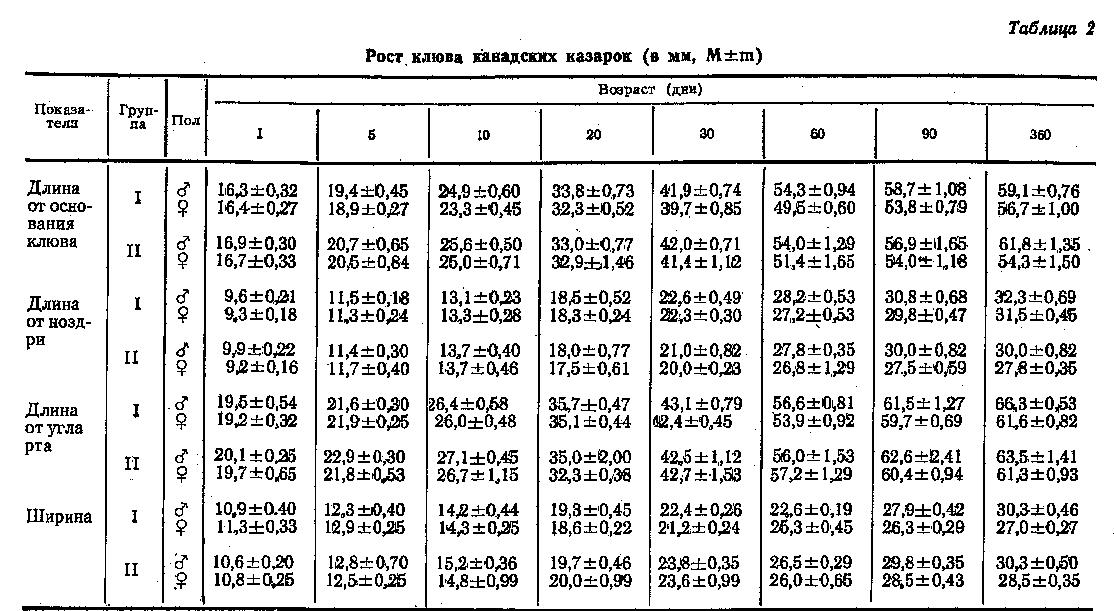
У суточных канадских казарок (как у самцов, так и у самок) в I и II группах длина клюва от основания составляла примерно 16 мм. Из данных, приведенных в таблице 2, видно, что увеличение клюва в длину у птиц проходило с равной скоростью. У годовалых канадских казарок различия в длине клюва между птицами I и II групп составило всего лишь 2,4-2,7 мм, причем у самцов преимущество было за птенцами II группы, а у самок - I группы. Однако статистически достоверных отличий не получено.
Длина клюва от ноздри у суточных птенцов I и II групп была одинаковой (табл. 2). Годовалые самцы I группы имели клюв на 2 мм длиннее, чем II. У самок обеих групп до 20-дневного возраста клюв в длину рос с одинаковой скоростью, далее - несколько быстрее у самок I группы, так что к 30 дням клюв у них стал на 2 мм длиннее, чем у птиц II группы. К 60 дням это различие нивелировалось, а затем проявилось снова и у годовалых птиц I группы клюв стал на 3,7 мм длиннее, чем у казарок II группы. Однако, эти отличия статистически не достоверны.
Та же картина прослеживалась и по длине клюва от угла рта. Рост клюва у самцов I и II групп был одинаков. У самок наблюдалось небольшое преимущество в росте у птиц II группы в период от месячного до 2-месячного возраста (различие в 2-месячном возрасте - 5 мм). Конечные результаты у годовалых птиц не имели статистически достоверных различий.
Из таблицы 2 видно, что ширина клюва у суточных казарок I и II групп (как у самцов, так и у самок) составляла II мм. В течение всего изученного периода постэмбриогенеза изменения размеров клюва в ширину у обоих групп были идентичны и у годовалых птиц клюв имел ширину 28-29 мм.
Для анализа возрастных изменений пропорций туловища канадских казарок измеряли длину и ширину туловища (табл. 3.). У только что вылупившихся птенцов длина туловища составляла 19-27%, а ширина – 18-20 % от размеров взрослых птиц. Поскольку рост туловища в длину вначале происходит интенсивнее, чем в ширину, у месячных птиц длина туловища достигала 68-78 %, а ширина - только 50-56 % соответствующих размеров взрослых птиц. К 2-месячному возрасту длина туловища выросла уже до 82-98 %, а ширина груди - до 83-92 %. У 3-месячных птиц эти размеры составили соответственно 90-100 и 90-98 %.
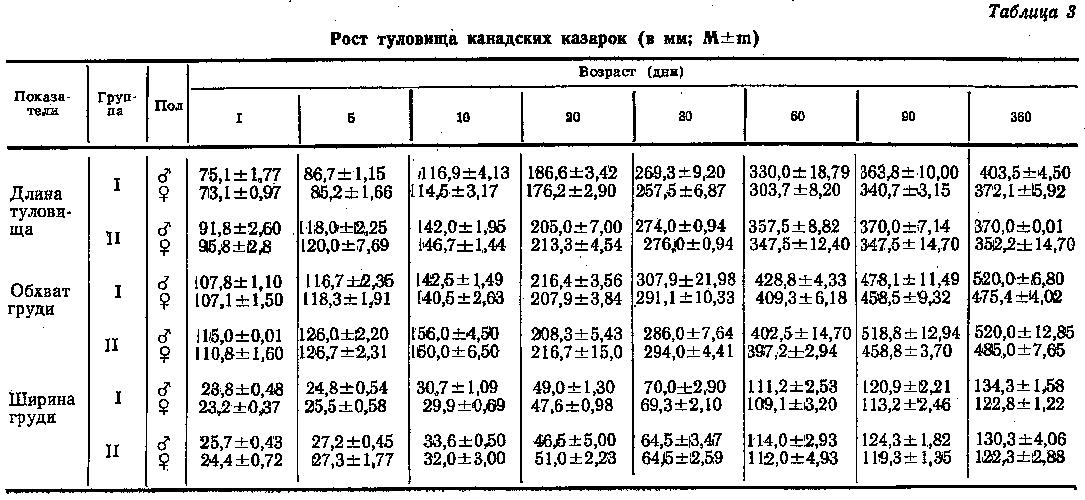
Суточные казарки I и II групп отличались по длине туловища. Так, у самцов II группы оно на 17 мм длиннее, чем у самцов I группы, а у самок это отличие было больше и составляло 23 мм. К месячному возрасту разница в длине туловища у самцов уменьшилась до 5 мм, а у самок - до 19 мм, а к 2-месячному возрасту разница в длине туловища у самцов уменьшилась до 5 мм, а у самок - до 19 мм, а к 2-месячному вновь увеличилась и составила у самцов 27 мм, а у самок - 45 мм. В дальнейшем у всех птиц I группы скорость роста туловища в длину возросла и к 90-му дню длина туловища птиц обеих групп почти сравнялась, а к году птицы I группы даже обогнали по этому показателю птиц II группы: самцы - на 33 мм, а самки - на 20 мм. Самцы и самки канадских казарок I группы по длине туловища статистически достоверно отличались от птиц II группы.
В ширину туловище росло у обеих групп казарок с одинаковой интенсивностью.
Развитие грудного пояса и изменение с возрастом формы передней части туловища характеризуется также величиной обхвата туловища (табл. 3). У суточных птиц обхват груди составлял 23 % от размеров взрослых птиц. К месячному возрасту обхват груди увеличился до 55-61 %, к двухмесячному - до 77-86 % и к трехмесячному - до 92-99 %.
Из табл. 3 видно, что обхват груди у самцов суточных канадских казарок I группы был на 7 мм, а у самок — на 4 мм больше, чем у птиц I группы. В первом месяце постэмбриогенеза рост груди в ширину у самцов обеих групп протекал с переменной интенсивностью и к концу месяца абсолютные показатели у птиц I группы были на 21 мм больше, чем у II группы. К двухмесячному возрасту это различие возросло до 27 мм. Однако к трем месяцам уже самцы II группы на 41 мм превосходили по обхвату груди птиц I группы, а к годовалому возрасту размеры их стали одинаковыми. У самок канадских казарок в течение первых двух месяцев жизни грудь в ширину интенсивнее росла у птиц I группы и в конце месяца она была на 12 мм шире, чем у казарок II группы. К трехмесячному возрасту обхват в области груди у самок обеих групп стал одинаковым, а к году этот показатель у птиц II группы стал на 10 мм больше, чем у птиц I группы. Однако различия статистически не достоверны.
Увеличение абсолютной длины крыла и предплечья казарок с возрастом показано в таблице 4. Длина крыла слагается из длины кисти и растущих маховых перьев. Темп роста крыла и предплечья неодинаков. Длина предплечья у суточных птиц составляла 12-13 %, а крыла лишь 7 % от их длины у взрослых птиц. Предплечье месячных казарок достигло 51-56 % окончательной длины, а крыло - всего лишь 20-25 %. Крыло развивается медленнее потому, что его длина зависит от роста не только кистевой части скелета, но и первостепенных маховых перьев, которые к этому времени еще не достаточно выросли. В течение второго месяца жизни канадских казарок происходил бурный рост перьевого покрова, так что длина крыла составляла уже 70-76 % от длины крыла взрослых птиц. Предплечье почти достигло размеров взрослых (98-99 %). У суточных самцов обеих групп канадских казарок крыло по длине не отличалось. До 2-х месячного возраста темп роста крыла у них был почти одинаков, однако затем крыло интенсивнее росло у птиц I группы и к годовалому возрасту было у них на 25 мм длиннее, чем у казарок II группы. Эта разница статистически достоверна.
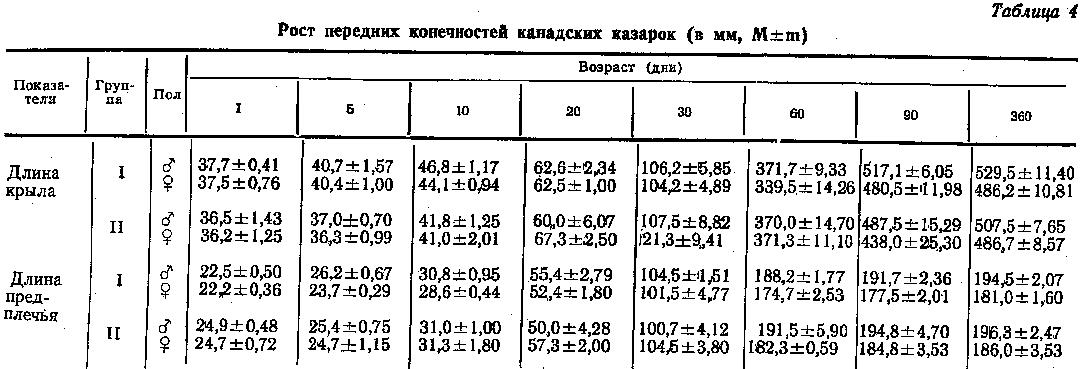
Из табл. 4 видно, что суточные самки обеих групп имели одинаковую длину крыла. В первые 2 месяца постэмбриогенеза интенсивнее росло крыло у птиц II группы, однако в дальнейшем темп его роста у птиц I группы увеличился и в результате у годовалых птиц длина крыла сравнялась.
У суточных казарок II группы (как у самцов, так и у самок) предплечье на 2 мм длиннее, чем у птиц I группы (табл. 4). В течение всего изученного периода развития рост предплечья у самцов обеих групп продолжался с переменной интенсивностью. У самок предплечье росло несколько интенсивнее у птиц II группы. Однако, к году его длина у самцов II группы была всего лишь на 2 мм, а у самок - на 5 мм больше, чем у птиц I группы. Это различие статистически не достоверно.
О возрастных изменениях длины цевки среднего пальца и когтя у канадских казарок можно судить по данным табл. 5. Все части задней конечности у суточных птиц хорошо развиты и их размеры достигают около 40 % размеров взрослых птиц. У месячных казарят ноги были самыми развитыми по сравнению со всеми изученными частями тела. Так, цевка достигала 89-96 %, а средний палец 84-92 % окончательной длины. Крыло в этом возрасте составляло лишь 21-24 % от окончательных размеров взрослых птиц. Это объясняется тем, что канадские казарки, как все водоплавающие, после вылупления легко перемещаются по суше и плавают, а крылья у них начинают функционировать значительно позже. У 2-месячных казарят крыло достигало 70-76 %, а цевка и средний палец уже почти заканчивали свой рост и составляли 95-99 % от их размеров у взрослых птиц.
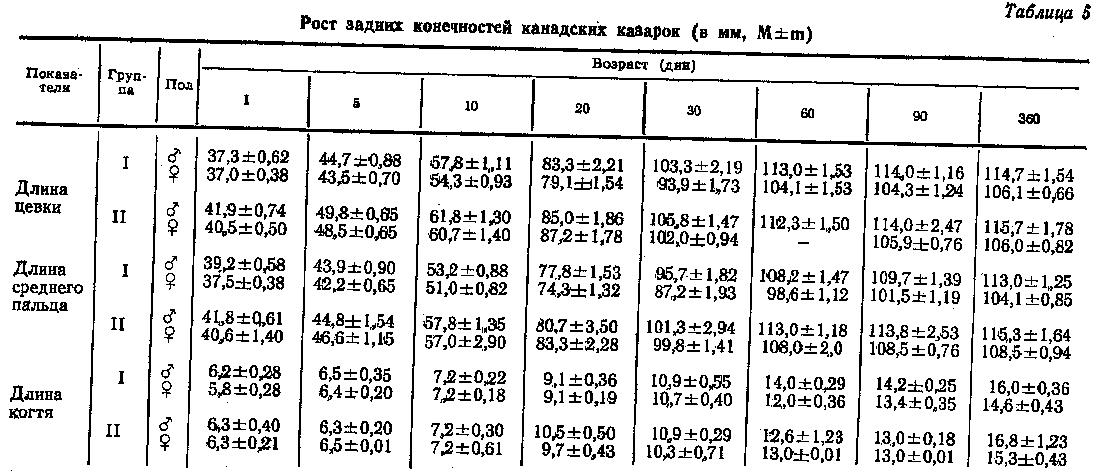
Цевка суточных казарок II группы на 3-4 мм длиннее цевки птиц I группы. У месячных самцов это различие составило всего 1,3 мм, а у самок - 7 мм; в дальнейшем оно все больше сокращалось и к двухмесячному возрасту исчезло.
Средний палец суточных казарят II группы на 1-2 мм длиннее, чем у птиц I группы. В течение первого месяца жизни птиц он более активно рос у II группы. Средний палец месячных самцов II группы на 3 мм, а самок на 8 мм длиннее, чем у I группы канадских казарок, и это преимущество сохраняется и в дальнейшем. У годовалых канадских казарок отличие в длине пальца составило 4 мм как у самцов, так и у самок, однако эта величина статистически не достоверна. Длина когтя среднего пальца зависит от условий содержания птиц (структуры поверхности пола) и полноценности кормления. Поскольку птицы I и II групп содержались в идентичных условиях, то и показатели роста когтя были одинаковые, что наглядно видно в табл. 5.
Проведенный эксперимент позволяет сделать следующие выводы:
1. При сравнении роста двух групп канадских казарок, которые выращивались в индентичных условиях, но получены от родителей разного происхождения, отмечены некоторые различия в интенсивности роста ряда частей тела на разных стадиях постэмбриогенеза.
2. Отмечавшиеся различия прослеживались, как правило, лишь от суточного до 90-днёвного возраста, но к году они исчезали или становились статистически недостоверными (за исключением длины туловища и длины крыла у самцов).
3. Анализ развития двух групп канадских казарок свидетельствует, что разработанная в ЦНИЛ технология содержания и кормления молодняка канадских казарок соответствует требованиям развивающегося организма и может быть использована в практике при разведении этого вида.