
Гражданкин А.В. Автономный контактный инкубатор и искусственная инкубация яиц диких птиц // Дичеразведение в охотничьем хозяйстве. Сборник научных трудов ЦНИЛ Главохоты РСФСР. Москва, 1985. С. 137-152.
При инкубации яиц диких птиц, предназначенных для разведения в специальных питомниках и зоопарках, а также во время их транспортировки из отдаленных районов нередко возникает необходимость в инкубаторе, который может работать без источников электроэнергии. Эта необходимость сохраняется и в том случае, если электроснабжение имеется, но возможны периодические перебои, вызывающие нарушение режимов инкубации. Во всех этих ситуациях не приходится рассчитывать на применение инкубаторов промышленных конструкций, в которых обогрев яиц и вентиляция воздуха происходят от внешних источников электроэнергии.
Конструкция автономного инкубатора должна учитывать также одно из основных требований - возможность воспроизведения в нем режимов инкубации, по всем параметрам максимально приближенных к естественным. В первую очередь необходимо, чтобы в яйцах можно было поддерживать вертикальный термоградиент требуемой величины в условиях неподвижного воздуха в инкубационной камере. При тесном взаимном расположении яиц в гнезде в присутствии насиживающей птицы среди яиц нет интенсивной вентиляции воздуха, нет иных источников увлажнения воздуха, кроме самих яиц (насиживающая птица увлажняет воздух в меньшей степени). Неизбежны при естественной инкубации и колебания температуры яиц, которые могут быть значительными. Тем не менее, в гнездах любого типа требуемый уровень аэрации яиц обеспечивается, а относительно перепадов температуры в ходе инкубации уже давно высказано соображение, что регулярные периодические охлаждения яиц необходимы для успешной инкубации (Третьяков, 1953; Рольник, 1968 и др.). Правда, но поводу вертикального термоградиента вопрос остается открытым. Несомненно, что он необходим, но в какой степени - неясно. При этом свойство белка и желтка менять свою плотность при изменении температуры способствует тому, что желток вместе с зародышем может вертикально перемещаться в яйце по термоградиенту, причем зародышу обеспечиваются требуемые температурные условия (Петров, 1981). Яйцо обладает, по сути, механизмом самонастройки температурного режима, осуществляющим «тонкую» подгонку при условии, что «грубую» обеспечивает насиживающая птица. Конструкция автономного инкубатора должна предусматривать возможность моделирования всех этих особенностей инкубационного режима с учетом видовых различий.
Наиболее богатый опыт инкубации яиц диких птиц накоплен для представителей отрядов пластинчатоклювых и курообразных. По другим отрядам опыт, как правило, невелик. В ряде случаев в инкубаторах с «куриным» режимом инкубации, а также под курами, утками и индейками выводятся птенцы самых разнообразных видов птиц. Однако вряд ли правомочно делать вывод о том, что «куриный» режим в равной мере пригоден для всех или большинства видов диких птиц, даже если в таких условиях были получены птенцы. Жизнеспособность молодняка при этом может оказаться пониженной.
Почти ничего не известно о том, насколько успешной может быть искусственная инкубация, если ее режим по основным параметрам будет соответствовать естественной, и какие отклонения допустимы, а также то, какие параметры являются определяющими при естественной инкубации. Эти вопросы требуют специальной экспериментальной проверки, в процессе которой те или иные параметры можно было бы поочередно изменять, а прочие поддерживать на естественном уровне, добиваясь в искусственных условиях максимальной имитации процесса естественной инкубации.
Исходя из вышеизложенного, мы сочли оптимальной такую конструкцию аппарата, при которой нагрев яиц будет производиться сверху путем непосредственного контакта с нагревателем. Идея такого прибора не нова. В промышленных масштабах она не получила широкого распространения в первую очередь, по-видимому, из-за сложности автоматизации регулировки процессов инкубации. Однако для моделирования естественных процессов при работе с небольшими партиями яиц аппараты такого типа, на наш взгляд, обладают большим преимуществом по сравнению с конвекционными и радиационными, что обеспечит соблюдение всех перечисленных требований.
Для работы был использован инкубатор оригинальной конструкции, в котором осуществляется контактный способ нагревания яиц, а воздух в инкубационной камере увлажняется за счет воды, испаряемой самими яйцами. Общее представление о конструкции можно получить по ее изображению на рисунке 1. Вверхней части в теплоизолирующем корпусе (3) размещается металлический резервуар из нержавеющей стали (5), имеющий в донной части 6 горловин, на которые надеты резиновые колпаки - «наседки» (6). Резервуар заполняется горячей водой. В нижней части (1) в теплоизолирующем корпусе установлена инкубационная камера из оргстекла (8), имеющая в донной части отверстия. В закрытом состоянии обе части теплоизолирующих корпусов скрепляются защелкой (9). Для подсветки во время операций, проводимых в темноте, к донной части нагревателя прикрепляется лампочка 2,5 В (7), питание которой осуществляется от трех элементов «Марс», размещенных в специальном блоке (2). Иного электропитания инкубатор не требует.
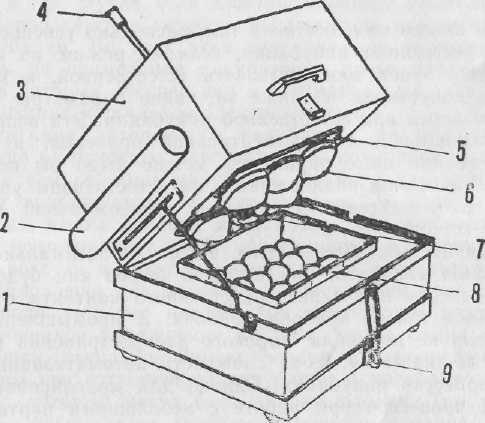
Рис. 1. Конструкция контактного инкубатора
1, 3 - теплоизолирующий корпус; 2 - блок питания; 4 - термометр; 5 - резервуар с горячей водой; 6 - «наседки»; 7 - электролампа подсветки; 8 - инкубационная камера; 9 – защёлка.
Наружные размеры корпуса инкубатора в собранном состоянии: длина 32 см, ширина 26 см, высота 28 см. Масса инкубатора, заполненного водой, 6,5 кг. Объем нагревательного резервуара 3 л. Размеры инкубационной камеры 21х14х6 см. Общая площадь вентиляционных отверстий в донной части инкубационной камеры 52 см2.
Яйца укладываются на подставки, положение которых регулируется таким образом, чтобы площадь зоны контакта яйца и «наседки» была 1,5-2 см2. Каждое яйцо контактирует с «наседкой» в своей верхней части со смещением к острому концу. При закрытом инкубаторе вентиляция происходит через щели в инкубационной камере. При открывании инкубатора для поворотов и взвешиваний яиц они вентилируются дополнительно.
Температура в верхней части инкубационной камеры регулируется периодической сменой воды в нагревателе, охлаждением яиц при открывании инкубатора и поворотом яиц. Контроль за температурным режимом производится при помощи ртутного термометра (4), помещенного в нагреватель, и двух термисторов типа CTЗ-17,касающихся скорлупы одного из яиц у его нижнего конца и в зоне контакта яйца с «наседкой». Заливка воды в нагреватель и ее слив производится через шланг, выведенный сбоку в нижней части корпуса (на рисунке не изображен).
Регуляция водного баланса яиц осуществляется подбором временных интервалов между разовыми охлаждениямии принудительной вентиляцией с таким расчетом, чтобы интенсивность влагоотдачи яиц в течение суток находилась на заданном уровне. При этом ориентация делается на предыдущие сутки. Контроль за динамикой влагоотдачи яиц осуществляется путем ежесуточного взвешивания каждого яйца на ручных весах с точностью до 0,01 г.
Данные о температуре окружающего воздуха записываются на ленте термографа, а затем на эту же ленту от руки наносятся показания термисторов, контактирующих с яйцом. При последующей обработке по температурным графикам мы рассчитывали среднечасовые уровни температуры, их вариации и т.п.
Поворот яиц па 180° вокруг оси производится вручную таким образом, чтобы наклеенная сбоку метка с номером яйца при повороте его проходила через верх.
Биологический контроль за эмбриональным развитием осуществляли ежесуточным овоскопированием каждого яйца, а с 8 суток инкубации еще и регистрацией движений яиц в воде.
Для работы были использованы 24 яйца обыкновенной пустельги. Из них 11 яиц (четные) составили контрольную группу, 11 - опытную. Из двух оставшихся одно оказалось неоплодотворенным, а другое было вскрыто на 13-е сутки инкубации для проверки общего хода инкубационного процесса. Контрольные яйца поворачивали 15 раз в сутки с 6 до 23 часов, а опытные (нечетные) - 5 раз: в 8, 11, 15, 18 и 20 часов. Общая программа инкубации представлена в таблице 1.
Таблица 1
Режимы инкубации яиц обыкновенном пустельги
(обозначения в тексте)
|
Параметр |
Литературные и расчётные (+) данные |
Источник информации |
Фактические данные при искусственной инкубации |
|
Среднечасовая Тзк |
38,6° |
Литвинов, Карабулин, 1979 |
38,3 ± 5,2° |
|
Допустимые колебания Тзкв течение суток |
37,6 - 40,1° |
Там же |
37,7 ± 39,0° |
|
Допустимые кратковременные изменения Тзк |
28,9-42,8° |
Там же |
28,5 ± 42,5° |
|
Вертикальный термоградиент в дни инкубации: 1-8 |
5 - 4° |
Там же |
5,0 ± 0,6° все дни |
|
9-13 |
3,5° |
Там же |
|
|
14-28 |
3,0° |
Там же |
|
|
Среднечасовая Тн |
35,0° |
Пономарёва, 1972 |
33,3 ± 6,2° |
|
Допустимые суточные изменения Тн |
33,3 – 36,8° |
Литвинов, Карабулин, 1978 |
32,1 – 34,3° |
|
Допустимые кратковременные изменения Тн |
22,6 - 38,4° |
Там же Пономарёва, 1972 |
27,0 - 37,0° |
|
Потеря массы яйца за весь период инкубации |
17 % от исходн. массы 15,0 ± 2,5 % от исходн., массы до 1-го проклева |
С. Бакаев, личное сообщение Ar, Rahn, 1978 |
14,3 % до проклева |
|
Продолжительность инкубационного периода |
27-29 суток |
Литвинов, 1983 |
26 суток до первого проклева |
|
Число среднесуточных поворотов каждого яйца |
15(+) |
Наши расчеты по данным Литвинова, Карабулина (1979) |
15 - контрольная группа, 5 - опытная группа |
Из контрольной группы вывелись все 11 птенцов, из опытной - 7 птенцов. 2 зародыша погибли на 4-е сутки инкубации, 1 - на 6-е, 1 - на 17-е сутки. Данные результаты были получены при условиях инкубации, описанных ниже.
ТЕМПЕРАТУРНЫЙ РЕЖИМ
Температура яиц в зоне контакта (Тзк) за весь периодинкубации поддерживалась на уровне +38,5±5,2 °С (X±Sx). В течение первой и последней перед вылуплением недель среднесуточный уровень Тзк равнялся +37,8 °С, а в течение второй и третьей недель +38,7 °С. Среднечасовые колебания Тзк, не считая специальных кратковременных охлаждений, невелики. Средний коэффициент вариации (CV) среднечасовой температуры в течение суток за весь период инкубации (27 суток) составил 2,8 ± 0,4 %.
Температура в нижней части яиц (Тн) в течение 4 недель инкубации была равна +33,3±6,2° (CV=1,8 %). Ее среднечасовые колебания, также без учета кратковременных специальных охлаждений яиц, в течение суток больше, чем для Тзк: коэффициент вариации 3,7±1,0 %. Вертикальный термоградиент яиц сохранялся па уровне 5,0±0,6°. В основном он определялся температурой нагревателя в зоне контакта с яйцами, интервалами между специальными охлаждениями яиц во время их поворотов и, в меньшей мере, температурой воздуха (Тв) около инкубатора, когда он закрыт. В то время, когда инкубатор находился в помещении и Тв в течение суток была наиболее стабильной (15-20°), коэффициент вариации Тв был равен 4-7 %. На открытом воздухе амплитуда колебаний Тв значительно повышалась: в ночное время воздух охлаждался до 4-12 °С, а днем нагревался до 30-38 °С. В таких условиях коэффициент вариации Тв достигал 30-50 %. Как видно из этих показателей, близкий к естественному температурный режим в инкубационной камере закрытого аппарата можно поддерживать независимо от внешних температурных колебаний.
При открывании инкубатора степень охлаждения яиц в основном определялась именно температурой воздуха вне инкубатора. Температура яиц на поверхности скорлупы в самые холодные периоды падала со скоростью до 1,2 град/мин, а при температуре воздуха 20 °С яйцо, нагретое в зоне контакта до 38 °С, остывало со скоростью до 0,4 град/мин.
Большие колебания температуры поверхности скорлупы в процессе инкубации не затрагивают более глубоких частей яйца. Предварительные опыты с яйцами серой вороны со сходной массой (около 19 г) показали, что при закрытом инкубаторе на поверхности скорлупы разница между температурой в верхней и нижней частях яйца стабилизируется па уровне 4,8-5,2° (инкубатор находится в помещении с Тв 18-20 °С), а непосредственно под скорлупой вертикальный термоградиенг яйца еще более стабилен и близок к 3,3°. Следовательно, колебания температуры яйца под скорлупой меньше, чем на ее поверхности. Одновременные измерения температуры в средней части яйца и на поверхности скорлупы показали, что воронье яйцо, нагретое в зоне контакта до 38 °С, при Тв 19 °С остывает за 10 минут на 5,4° на поверхности скорлупы и на 2,5° в центре. Вполне очевидно, что эмбрион не испытывает столь больших колебаний температуры, как окружающие его оболочки (белковая, скорлуповая). В то же время при закрытом инкубаторе на поверхности скорлупы температура практически такая же, как и в центре яйца. Для яиц пустельги за весь период инкубации она была равной 35,8°. Этому соответствовали показания ртутного термометра, помещенного в начале инкубации в инкубационную камеру на уровне середины лежащего на боку яйца. Впоследствии этот термометр был удален за ненадобностью, т.к. измерявшуюся им температуру, как было сказано выше, легко рассчитать по данным вертикального термоградиента.
В естественных условиях величина вертикального термоградиента яиц пустельги меньше, чем удавалось нам воссоздать в инкубаторе, при том, что температура в зоне контакта яйца с «наседкой» в инкубаторе и с насиживающей птицей в гнезде была одинаковой. Вполне очевидно, что в данном случае не только масса и размеры яиц могут определять величину вертикального термоградиента, но и характер субстрата, на котором они лежат. Подстилка естественного гнезда аккумулирует тепло яиц и воздуха и тем самым способствует повышению температуры нижней части яиц и снижению уровня вертикального термоградиента. В инкубаторе яйца находились на подставках и охлаждались поступающим к ним снизу через вентиляционные отверстия воздухом, что увеличивало вертикальный термоградиент. Данные факты необходимо учитывать при моделировании инкубационного процесса в том или ином типе инкубатора.
При определении естественного температурного режима инкубации немаловажно, в какой точке производятся измерения.
Наиболее просто измерять температуру в гнезде на дне лотка, т.к. термодатчик легко может быть замаскирован и не будет беспокоить птицу. Но при отсутствии данных о вертикальном термоградиенте яиц в конкретных условиях трудно судить об уровне температуры в середине яйца.
Наиболее сложно измерять температуру яиц на верхнем уровне, если только не вводить термодатчик в одно из яиц. Торчащий из гнезда датчик нередко вызывает тревогу у птиц, и они бросают гнездо (особенно часто это происходит в начале инкубации) или пытаются вырвать датчик из гнезда. Если птица не обеспокоена датчиком, то остается большая вероятность касания ее и датчика. При этом, естественно, возникнут ошибки в оценке температуры яиц. Возможность больших искажений информации в таких случаях велика.
На наш взгляд, наиболее точные показатели температурного режима можно получить при измерении температуры поверхности одного из яиц микротермистором,расположенным на уровне середины яйца, и, одновременно, другим термистором - на дне лотка. Если второе измерение сопряжено с техническими трудностями, можно ограничиться первым, а величину вертикального термоградиента яиц данного размера в конкретных условиях воссоздать в экспериментальных условиях на модельном яйце. Ошибки, возникающие при таком способе расчетов температурного режима, сводятся к минимуму. Насколько они могут компенсироваться действием механизма самонастройки яиц, о котором говорилось выше, будет ясно только после практического решения вопроса.
Для имитации слетов птицы с гнезда мы периодически охлаждали инкубируемые яйца во время их поворотов, оставляя инкубатор открытым на 2-15 мин. Наиболее продолжительные охлаждения (25-30 мин.) сопровождали процесс взвешивания. При температуре воздуха ниже + 15 °С после изъятия очередного яйца для взвешивания инкубатор закрывали во избежание излишнего охлаждения яиц. В общем продолжительность охлаждений определялась не столько необходимостью регуляции температурного режима, сколько была связана с задачей регуляции водного обмена, о чем будет сказано в дальнейшем. Независимо от продолжительности разовых охлаждений (суммарно 1,5-2,5 часа в сутки) среднесуточная температура яиц в зоне контакта и, тем более, в нижней части яиц менялась мало. Лишь в условиях пониженной температуры внешнего воздуха длительность разовых охлаждений яиц ограничивали в зависимости от степени их охлаждения: нижний предел охлаждения, в соответствии с программой, равен + 28,5 °С.
При повышенной внешней температуре охлаждение яиц при открытом инкубаторе, естественно, было незначительным. В общей сложности кратковременные колебания Тзк за всевремя инкубации не выходили за пределы 28,5-42,5 °С. Предельные значения температуры сохранялись не более 10-15 мин., соответственно, в холодную погоду при взвешивании и при заливке горячей воды в нагреватель. При закрытом инкубаторе среднечасовые колебания Тзк не выходили за пределы 37,7-39,0 °С. В нижней части яиц кратковременные колебания температуры наблюдались в пределах 27-37 °С, а среднечасовые в течение суток - 32,1-34,3°.
ВОДНЫЙ РЕЖИМ И РЕГУЛЯЦИЯ ВОДНОГО БАЛАНСА ЯИЦ
Технические сложности прямого измерения влажности воздуха в инкубационной камере не позволили измерить этот параметр непосредственно. Однако, используя имеющиеся данные по водопроницаемости скорлупы и среднесуточной усушке яиц пустельги, мы можем дать ему оценку.
По литературным данным, согласно измерениям, проведенным на двух яйцах пустельги, водопроницаемость скорлупы (G) равна 3,98 мг/сутки х торр (Ar, Rahn, 1978) или 2,98 мг/сутки х гПа. По нашим измерениям на коллекционном материале из Зоологического музея МГУ этот показатель для 26 ненасиженных яиц равен 6,0±2,2 мг/сутки х гПа. Наши данные получены с использосанием гигрометра и психрометрического метода (Гражданкин, 1981). По уравнению, связывающему водопроницаемость скорлупы с суточными влагопотерями яйца (МH2O,мг), G = MH2O /Δ ДПВ, где Δ ДПВ - градиент давления паров воды через скорлупу в гПа, можно рассчитать ДПВ вне яйца, т.е. в воздухе инкубационной камеры. В условиях наших опытов суточные влагопотери яиц составили 132±29 мг, следовательно, градиент ДПВ равен (132:6)=22 гПа. В яйце ДПВ соответствует давлению насыщенного водяного пара (Гражданкин, 1983) и при средней температуре инкубации +35,8 °С оно равно 58,8 гПа. Следовательно, ДПВ вне яйца равно (58,8-22)=36,8 гПа, что при данной температуре воздуха соответствует относительной влажности воздуха 63%.
Измерения относительной влажности воздуха вне инкубатора аспирационным психрометром показали, что воздух здесь намного суше, чем в инкубаторе: в помещении ДПВ равно 12,4 гПа, вне помещения - около 6 гПа. Вполне очевидно, что сами яйца достаточно интенсивно увлажняют воздух вокруг себя и дополнительных увлажнителей в инкубаторе данного типа не требуется. Общее время ежесуточного пребывания яиц на открытом воздухе составило в среднем 88±23 мин, (от 45 до 168 мин). Даже такое продолжительное воздействие сухого воздуха не вызвало чрезмерного усыхания яиц. Среднесуточные влагопотери яиц соответствовали естественному уровню, известному из литературных источников (табл. 1,2).
Таблица 2
Динамика суточных влагопотерь яиц обыкновенной пустельги при искусственной инкубации
|
№ кладки |
№ яйца |
Суточные влагопотери (мг) в дни инкубации (Х±Sx) |
||
|
1-7 |
8-14 |
15-20 |
||
|
I |
1 |
110±22 |
118±18 |
107±12 |
|
2 |
97±20 |
110±20 |
113+11 |
|
|
3 |
89±8 |
108±20 |
109±14 |
|
|
4 |
107±19 |
109±14 |
119±19 |
|
|
II |
5 |
94±12 |
107±18 |
проклев |
|
6 |
98±9 |
106±16 |
проклев |
|
|
7 |
89±18 |
98±34 |
проклев |
|
|
8 |
110±31 |
126±13 |
проклев |
|
|
III |
10 |
136±27 |
133±25 |
126±26 |
|
12 |
156±21 |
152±9 |
150±8 |
|
Взвешивание яиц показало, что даже в одной кладке есть яйца с существенно различающимися уровнями водопроницаемости скорлупы, вследствие чего суточные влагопотери таких яиц различались в 1,6-2,4 раза. Определенную роль в изменениях скорости влагоотдачи играл индивидуальный микроклимат около каждого яйца и его изменчивость. Об этом говорит тот факт, что динамика суточных влагопотерь яиц не была синхронной. Для инкубаторов с отсутствием специальной вентиляции микроклимат может оказывать большое влияние на регуляцию водного обмена яиц, поэтому данному вопросу следует уделить внимание.
Как известно, уровень водопроницаемости скорлупы определяет толщину пограничного со скорлупой слоя воздуха, его водопроницаемость и влияет на интенсивность испарительной влагоотдачи яиц (Spotila et al., 1981; Гражданкин, 1983). Пользуясь этим свойством, мы провели эксперимент, целью которого была попытка направленного воздействия на индивидуальные режимы водного обмена у разнокачественных яиц.
Из 24 яиц мы составили 6 групп, объединив в каждой по 4 яйца с близкими уровнями суточных влагопотерь в один из дней в середине инкубационного периода. Средние суточные влагопотери непосредственно перед экспериментом (14 мая) составили для I-VI групп, соответственно, 57, 76, 86, 94, 122 и 166 мг. Группы яиц разместили в инкубаторе таким образом, чтобы яйца с наименьшей водопроницаемостью скорлупы и наименьшей суточной усушкой находились в одном конце, а яйца с повышенной водопроницаемостью скорлупы (группы V и VI) - в другом. Сбоку в инкубационную камеру был вставлен небольшой вентилятор так, чтобы ближе всего к нему находились яйца I и II групп. Помимо обычной вентиляции при открывании инкубатора воздух в инкубационной камере при закрытом инкубаторе обновляли 8 раз в сутки, включая вентилятор на 1-3 с в перерывах между поворотами яиц.
Как и следовало ожидать, испарительная влагоотдача яиц в целом увеличилась. 15 мая она составила для I-VI групп, соответственно, 102, 107, 117, 110, 125 и 150 мг. Вместе с этим снизился индивидуальный разброс уровней влагоотдачи за счет того, что испарение воды из яиц вблизи вентилятора увеличилось сильнее, чем у отдаленных яиц. Если 14 мая коэффициент вариации влагопотерь составил 37 %, то 15 мая он был 26 %. Некоторое снижение уровня влагопотерь 15 мая по сравнению с предыдущим днем у яиц VI группы произошло, по-видимому, из-за большей влажности воздуха вне инкубатора в этот день (этот параметр мы не измеряли).
16 мая вентилятор был убран, и соотношение влагопотерь в группах стало следующим: 67, 100, 102, 90, 105 и 163 мг. Проявилась четкая тенденция к увеличению влагопотерь у яиц с пониженной водопроницаемостью скорлупы по сравнению с влагопотерями 14 мая. Контрольная проверка взвешиванием 17 мая подтвердила наличие этой тенденции. Хотя общий уровень испарительной влагоотдачи яиц повысился, видимо, из-за снижения влажности внешнего воздуха по сравнению с предыдущим днем, тенденция к увеличению влагопотерь у яиц с пониженной водопроницаемостью скорлупы сохранилась. Соотношение влагопотерь в I-VI группах стало следующим: 92, 113, 132, 120, 137 и 180 мг.
Достигнутый эффект мы объясняем следующим образом. При смешанном расположении разнокачественных яиц те, что обладают большей водопроницаемостью, испаряют больше, воды в окружающий воздух и подавляют испарительную влагоотдачу соседних яиц с пониженной водопроницаемостью скорлупы, т.к. повышают влажность и, соответственно, снижают водопроводимость пограничного со скорлупой этих яиц воздуха. Чем больше различия водопроницаемости скорлупы у яиц, тем больше и воздействие, имеющее практически односторонний характер. Совместная группировка яиц с минимальной водопроницаемостью скорлупы приводит к тому, что суммарная влагоотдача группы снижается, воздух увлажняется меньше, и это способствует усилению влагоотдачи каждого отдельно взятого яйца в группе. К тому же снижается до минимума подавляющее влагоотдачу влияние отдельных яиц на соседние, и интенсивность индивидуальной влагоотдачи выравнивается. Как следствие таких изменений — наблюдаемое в эксперименте повышение интенсивности влагоотдачи яиц с низкой водопроницаемостью скорлупы, когда они находятся в группе с себе подобными, и уменьшение индивидуального разброса величин влагопотерь.
Как мы и предполагали, группировка яиц по характеруводопроницаемости скорлупы и их расположение относительно вентилятора привели к усилению скорости влагоотдачи яиц с пониженной водопроницаемостью скорлупы и приблизили ее к таковой других яиц. Но полного выравнивания уровней влагопотерь мы не достигли. По всей вероятности, одностороннее подавляющее воздействие одних яиц на другие недостаточно велико, а вместе с тем картина динамики испарительной влагоотдачи яиц смазывается периодическими открываниями инкубатора, разрушающими пограничные слои воздуха у скорлупы. Вдобавок ко всему, проявление эффекта нарушается еще и из-за непостоянства влажности внешнего воздуха, поступающего в инкубатор.
Проведенные эксперименты уже в первом приближении показали, что простой перегруппировкой яиц можно добиться некоторого увеличения скорости испарения у яиц с низкой водопроницаемостью скорлупы, но снизить уровни влагоотдачи у яиц с повышенной водопроницаемостью скорлупы при этом не удается. Сочетание перегруппировки яиц с применением вентилятора (размещение групп яиц в градиенте воздушного потока) способно привести к повышению влагоотдачи яиц в разной мере с одновременным снижением индивидуальных различий. Но при таком способе регуляции водного режима яйца с высокой водопроницаемостью скорлупы следует изолировать от вентилятора во избежание их пересыхания.
В некоторых случаях при работе с инкубаторами с неподвижным воздухом к группировке яиц для регуляции их водного баланса можно прибегнуть, если приходится иметь дело с большой партией яиц, имеющих существенно разнокачественную скорлупу. Мы этого не делали по той причине, что в нашем инкубаторе яйца испаряли воду в пределах допустимого. На результатах инкубации не сказались отрицательно ни минимальные уровни усушки яиц (0,41 % от исходной массы яйца в сутки), ни максимальные (0,9 % х сутки-1). По окончании опыта яйца были возвращены на прежние места в инкубационной камере.
Суточные влагопотери яиц пустельги составили в среднем 0,64±0,14 % от исходной массы (n=17) в течение 13-22 суток инкубации. В пересчете на 26 суток (продолжительность инкубации до первого наклева) это должно привести к потере массы 14,3 %, что близко к расчетному уровню для яиц с аналогичной исходной массой (21 г). Немедленно после проклева скорость испарительной влагоотдачи повышается, достигая 530±150 мг/сутки (n=7), т.е. увеличиваясь в среднем в 4 раза по сравнению с обычной скоростью влагоотдачи. Мы не смогли проследить увеличения скорости испарительной влагоотдачи, которое можно было бы связать с изменениями водопроницаемости скорлупы в ходе инкубации. В одних случаях отмечена тенденция к незначительному повышению уровня влагоотдачик концу инкубационного срока, в других, наоборот, отмечалась тенденция к снижению уровня влагопотерь (табл. 2). Вполне возможно, что такая разница в интенсивности усушки яиц, находящихся в одной камере, обусловливается различиями микроклимата около яиц, о чем говорилось выше.
Хотя основная часть воды испаряется из яиц до появления. Наклевов, после них доля влагопотерь от суммарных была значительной. Особенно она возрастала, если интервал между первым проклевом и моментом выведения птенца увеличивался. Несмотря на то, что мы не предпринимали никаких мер для повышения влажности воздуха в инкубационной камере в период вылупления птенцов, как это рекомендуется для инкубаторов с принудительной вентиляцией, никаких отрицательных последствий этого у птенцов мы не отмечали. Все птенцы выводились самостоятельно, хорошо обсыхали в выводковой камере и имели нормально втянутую пуповину.
ПОВОРОТЫ ЯИЦ
Уловить заметную разницу в развитии аллантоиса у яиц опытной и контрольной групп (напомним, что в опытной группе яйца поворачивали реже) мы не смогли. Сложность наблюдений за эмбриональным развитием усугублялась темной, пятнистой окраской скорлупы, затрудняющей овоскопирование, - а также тем, что яйца находились на разных стадиях развития. Поэтому у нас нет оснований связывать смертность эмбрионов из опытной группы (4 из 11) с ненормальным развитием аллантоиса. Однако, можно предположить, что для свежеотложенных яиц опытной группы температура в первые дни инкубации могла оказаться слишком высокой по сравнению с естественной, так как нормальная температура инкубации эмбрионов на поздних стадиях развития выше, чем на начальных. Уменьшение числа поворотов яиц в этой ситуации могло способствовать некоторому перегреву зародышевых дисков и в результате вызвать присыхание зародышей к подскорлуповым оболочкам. Эти признаки были обнаружены нами у зародышей в яйцах № 9 и № 23 на 4-е сутки инкубации и у № 11 - на 6-е сутки. Причины гибели одного зародыша на 17-е сутки инкубации (№ 17) остались неясными. Возможно, сказалось отрицательное воздействие повышенной водопроницаемости скорлупы и избыточной усушки яйца. Среднесуточные влагопотери этого яйца были максимальными и составляли 198 мг (на 69 % выше среднего уровня для всех яиц).
Успешная инкубация яиц при соблюдении режимов, заданных программой для контрольной группы, свидетельствует о том, что данная программа может быть принята за оптимальную. В течение всего срока инкубации приемлемы одинаковые режимы температуры, влажности воздуха и числа поворотовяиц. Однако для первых 2-3 суток и после проклева, возможно, температуру в зоне контакта следует поддерживать не выше 37 °С. При этом яйца необходимо поворачивать не менее 10 раз в сутки. Температура в зоне контакта может быть уменьшена и за счет уменьшения площади контакта «наседки» с яйцами опусканием подставки. В нашем инкубаторе после появления проклевов мы снимали яйца с подставок и укладывали их на дно инкубационной камеры на бумажно-полиэтиленовые прокладки. В таком положении яйца лишь слегка касались «наседок», которые уже не создавали помех при вылуплении птенцов.
Мы убедились, что в инкубаторе с контактным нагревом яиц и небольшим объемом инкубационной камеры легко могут быть воспроизведены режимы температуры яиц и влажности воздуха, соответствующие естественным. При этом температурный режим достаточно контролировать лишь по температуре в зоне контакта, если есть опыт работы с конкретной конструкцией инкубатора и с яйцами известного размера. Контроль за режимом влажности воздуха можно и достаточно осуществлять по ежесуточной динамике изменений массы яиц. При работе с большой партией яиц можно взвешивать не все яйца, а лишь те, что имеют крайние значения водопроницаемости скорлупы, которые пропорциональны величине усушки яиц. Это устанавливается в первые двое-трое суток работы взвешиванием всех яиц.
Достаточный уровень газообмена в данном инкубаторе подтверждается не только нормальным ходом влагоотдачи яиц, но и тем, что не было гибели эмбрионов даже в тех случаях, когда в инкубаторе одновременно находились 5-6 проклюнутых яиц, а инкубатор оставался закрытым в течение 4-6 часов (ночное время). В эти периоды газообмен осуществлялся только путем пассивной диффузии через вентиляционные отверстия.
Приведем теперь некоторые биологические характеристики яиц, использовавшихся для инкубации.
Размеры яиц (n=23). Длина 40,1±1,3 мм, диаметр 31,3+0,9 мм (X±Sх).
Масса яиц. Взвешивание свежеотложенных яиц не представлялось возможным, поэтому следует говорить лишь о восстановленной массе, рассчитывая испарительные потери массы по среднесуточным влагопотерям за период инкубации и продолжительности инкубационного периода и суммируя потери массы с массой яйца в конце инкубации. Для 20 яиц среднесуточные потери массы составили 117±28 мг. Масса свежеотложенного яйца пустельги составила в среднем 21,20±1,71 г. Следовательно, ежесуточная усушка составила в среднем 0,55±0,13 % от массы свежеотложенного яйца (0,64±0,14 % от массы яйца в момент закладки в инкубатор). Динамика суточных влагопотерь нестабильна, как это было сказано выше, поэтому точность расчетов восстановленной массы снижается по мере увеличения временного интервала между откладкой яйца и его первым взвешиванием. Однако если приходится иметь дело с малонасиженными яйцами, расчеты допустимого уровня влагоотдачи можно производить, исходя из величин массы яиц в день закладки их в инкубатор.
Биологический контроль. Границы аллантоиса просматриваются в яйцах пустельги более или менее удовлетворительно с начальных стадий до его смыкания в остром конце яйца. Видны лишь самые крупные кровеносные сосуды, а мелкие маскируются пятнистостью скорлупы. Движения эмбрионов выявляются при овоскопировании в первой половине инкубационного срока, а позднее - при опускании яиц в воду. Первые вращательные движения яиц в воде мы отметили на 8-13-е сутки инкубации. В основном появление этих движений совпадало с моментом, когда яйцо стало всплывать. Но у некоторых яиц с низкой водопроницаемостью скорлупы первые движения при погружении в воду отмечались тогда, когда они еще невсплывали.
В конце первой декады инкубационного периода, когда движения яиц в воде только появлялись, они были нерегулярными и иногда их не удавалось уловить в течение 1-3 мин. В нескольких случаях мы не отмечали движений яиц в воде за время, отведенное биологическому контролю, в течение 2 суток, поэтому создавалось впечатление гибели зародышей уже после того, как у них были отмечены первые движения.
Проклев птенцов. Во всех, за исключением одного, случаях зародыши правильно располагались в яйцах, и первый наклев появлялся ближе к тупому концу. Расположение наклева относительно зоны контакта с «наседкой» не было фиксированным. Все яйца за 1-2 суток до расчетного дня проклева переставали поворачивать. Не замечено приуроченности моментов появления наклевов к определенному времени суток. Последующие наклевы появлялись примерно через сутки после первого. Чаще они располагались около первого, реже - в ином месте.
С момента наклева до начала активного вылупления проходило 39-60 часов. За это время птенцы более или менее часто подавали из яиц громкие звуковые сигналы в виде писка. Частота вокализации была индивидуальной, нестабильной, но в ночное время птенцы обычно замолкали. Время, затрачиваемое птенцами на активное вылупление, составляло 15-40 мин. Вылупление птенцов всегда происходило в светлое время суток - от 6 до 16 часов поясного времени.
Постэмбриональное развитие. Масса 7 птенцов, измереннаясразу после вылупления, была 13,39±0,50 г (12,75-14,10 г).Скорлупа с остатками оболочек и экскрементами весила2,43±0,09 г. Средняя масса новорожденных птенцов составила67,4±2,3 % от массы свежеотложенных яиц.
Птенцы в день вылупления или через 2-3 суток возвращались нами в те же гнезда, откуда были взяты яйца. Наблюдения за их развитием показали, что оно проходило нормально и не отличалось от такового других птенцов, выведенных пустельгами. Убедившись, что в инкубаторе птенцы выводятся нормально, в дальнейшем мы возвращали в гнезда яйца с появлением первого наклева. Это снижало беспокойство птиц, еще не имевших в гнездах собственных птенцов. Подкладывание птенцов в гнезда с яйцами вызывало у взрослых птиц отрицательную реакцию. Одна из самок, вернувшись к гнезду после подкладки ей птенцов, тут же улетела и не возвращалась в течение 40 минут. Во избежание переохлаждения птенцов и яиц мы изъяли птенцов и вернули их снова через 2 дня, когда в гнезде уже появился один собственный птенец. В этом случае самка быстро вернулась к гнезду и приняла подложенных птенцов.
Эти факты необходимо иметь в виду, если ставится задача искусственной инкубации яиц хищных птиц с последующим возвращением птенцов родителям. Возвращение яиц с проклевами насиживающие птицы воспринимают спокойно и в том случае, если в гнездах находятся птенцы. Самки продолжают насиживать эти яйца.
ЛИТЕРАТУРА
Гражданкин А.В. Видовые различия водопроницаемости скорлупы яиц мелких воробьиных птиц, определяемые при помощи гигрометра. - Зоол. журн., 1981, т. 60. вып. II, с. 1704-1711.
Гражданкин А.В. Влияние влажного воздуха на испарительную влагоотдачу птичьих яиц. - Зоол. журн., 1983, т. 62, вып. 4, с. 585-591.
Литвинов Н.А., Карабулин В.В. Температура инкубации кладки у обыкновенной пустельги. - В кн.: Гнездовая жизнь птиц. Пермь, 1979. с. 20-23.
Петров Б.К. Терморегуляторные механизмы яиц некоторых видов птиц и их значение в процессах насиживания. - Автореф. дис. ... канд. биол. наук. М., 1981. 16 с.
Пономарева Т.С. Температурные условия в гнездах некоторых видов птиц аридных территорий. - Зоол. журн., 1972, т. 51, вып. 12, с. 1846-1856.
Рольник В.В. Биология эмбрионального развития птиц. Л.: наука, 1968. 424 с.
Третьяков Н.П. Инкубация. - М.: Гос. изд-во сельскохоз. литературы, 1953, 312 с.