
Иванова В.С., Трошкина Н.Н., Канакова А.В. Постэмбриогенез кряквы при искусственном разведении // Разведение ценных и редких видов животных. Сборник научных трудов ЦНИЛ Главохоты РСФСР. Москва, 1987. С. 82-96.
Получать на дичеферме максимальное количество продукции можно только при правильном выращивании молодняка, при знании закономерностей индивидуального развития птиц и факторов, влияющих на него.
Рост и развитие крякв изучали на примере птиц, выращенных в питомнике ЦНИЛ на режиме, разработанном в отделе дичеразведения для промышленного разведения кряковых уток. Наблюдения проводили за изменением массы тела и экстерьера птиц в возрасте 1, 5, 10, 20, 30, 60, 90, 120 дней.
Утята были выведены из яиц крякв, содержащихся в питомнике ЦНИЛ. Из инкубатора утят сразу же помещали по 15-20 голов в трехъярусные клетки (42х32x21 см) с обогревом от лампы накаливания (15-25 Вт). Сетчатый пол застилали мешковиной, ставили воду и корм. По мере обсыхания утят пересаживали в металлические сетчатые трехъярусные клетки размером 110x65 см и высотой 35 см каждая. Ячея сетчатого пола 1,25х1,25 см. Первые 5-7 дней, чтобы у утят не проваливались ноги, весь пол застилали мешковиной, а далее - лишь небольшой участок под грелкой. Под сетчатым полом находился выдвижной поддон для сбора помета и остатков корма. Утята во время питья сильно расплескивали воду, поэтому под поилкой на поддон ставили кювет для сбора воды. Инфракрасные лампы для обогрева утят располагали так, чтобы они обогревали только половину площади пола. Кормушки и вакуумные поилки ставили в противоположной стороне от обогревателя. Такие клетки были размещены в помещении брудерной комнаты.
При включенных обогревателях и температуре в помещении 20-25 °С внутри клеток температура воздуха была 28-30 °С, а под грелками - до 35 °С. По мере роста утят источники обогрева поднимали (или отодвигали) для уменьшения нагрева: во вторую пятидневку до 22-26 °С, затем - до 18-20 °С. Относительную влажность воздуха стремились поддерживать, на уровне 60 %.
В помещении брудерной молодняк содержали до 10-15 дней. Отход утят наблюдался с 4-го по 5-й день и состоял только из слабых птиц. В дальнейшем падеж был незначительным, и сохранность молодняка составляла 95-85 % за все время выращивания.
Из брудерной утят переводили в акклиматизатор, переходя к напольному содержанию. Акклиматизатор состоял из деревянного закрытого помещения размером 2x2 м, пристроенного к нему навеса такой же площади и сетчатого выгула 2х8 м. Высота помещений и навеса 2-2,5 м, выгула - 1,5 м. Пол в закрытом помещении деревянный, покрытый линолеумом, на который насыпан слой сухого, песка. Здесь же установлен обогреватель, который нужен птенцам до 3-недельного возраста, если температура воздуха ниже 15 °С.
В первые 2 дня после перевода в акклиматизатор утят содержали, только в закрытом помещении. При теплой, сухой погоде на 3-4-й день утят выпускали под навес (в 10-12 чдня). Навес от выгула отделяли сетчатым щитом высотой 50 см. Через 2 дня (если не было дождя) утят выпускали в сетчатый выгул, где был установлен бассейн размером 1,2х1,2 м и глубиной 0,3 м. Приэтом следили, чтобы утята излишне не измокали, так как копчиковая железа у них еще недостаточно функционирует. В акклиматизаторе птиц содержали до 120 дней, а далее переводили в родительское стадо.
Изменения массы тела и относительных среднесуточных привесов молодняка в постэмбриогенезе отражены на рис. 1. Полученные данные позволили проанализировать изменение весового роста кряков в течение постэмбрионального периода. Нарастание массы тела утят происходило непрерывно, но неравномерно, до 90-дневного возраста, после чего, в основном, завершалось. До 30 дней масса тела селезней и уток возрастала с одинаковой скоростью, затем рост селезней стал опережать рост уток.
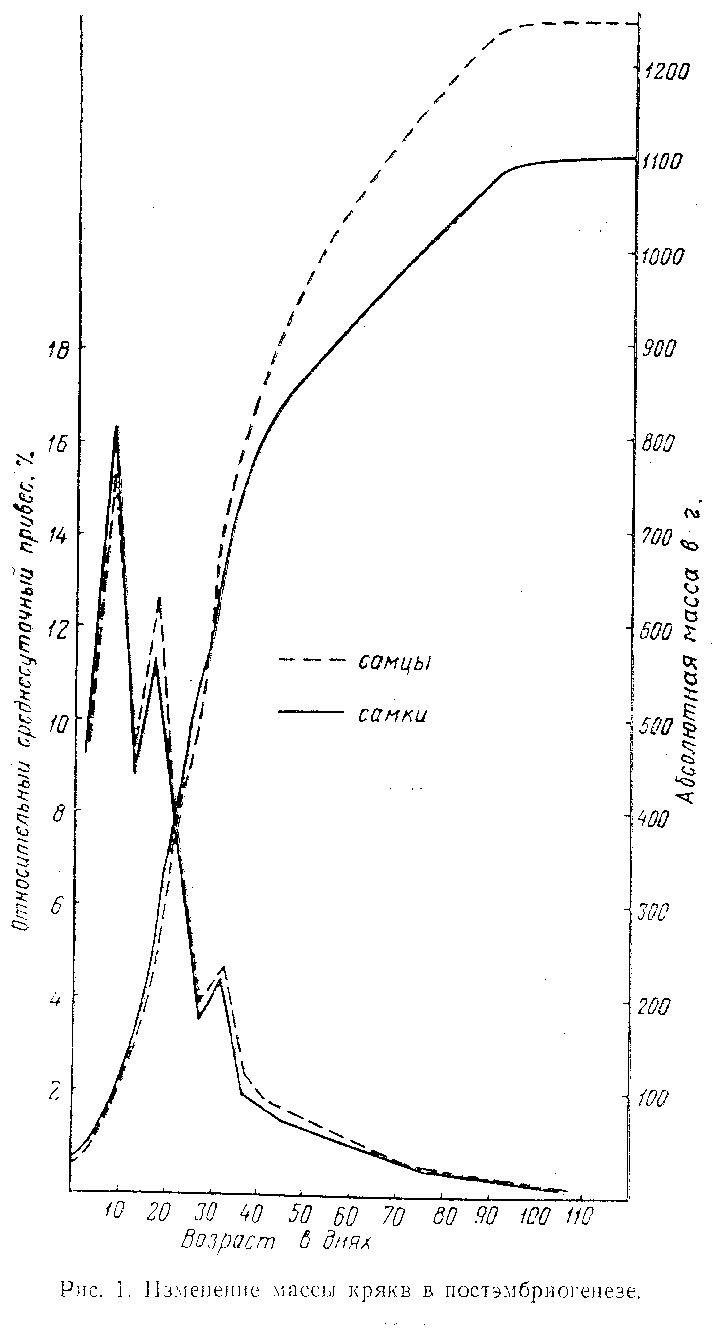
Кривая относительной скорости роста массы тела с момента вывода утят стремительно шла вверх, достигая на 10-й день максимального подъема (у селезней около 15 % и уток более 16 %). С 10-го дня темп роста снижался до 9 %, что связано с началом роста перьевого покрова, а с 15 по 20-й день вновь увеличивался до 12,5 %, образуя второй пик. Далее уровень прироста до 25-го дня резко снижался, составляя около 4 %, а с 25 по 30-й день вновь увеличивался, образуя 3-ю волну. После 30 дней кривая приростов постепенно приближалась к оси абсцисс (рис. 1).
Степень увеличения массы тела утят за период эмбриогенеза (отношение массы только что вылупившегося утенка к массе взрослой птицы) составляла у селезней 2 % ау уток 3 %. В первый месяц постэмбриогенеза утята быстро росли и к концу его достигали половины массы взрослых крякв. Масса уток составляла 98 % массы селезней.
Масса тела двухмесячных птиц достигала уже 85 % массы взрослых. Селезни весили 1068 г, утки - 940 г, или 88 % массы селезней — появились признаки полового диморфизма по массе тела (Р=0,002).
В трехмесячном возрасте весовой рост утят в основном завершен, так как селезни достигали массы взрослых (1250 г), а масса уток составляла 99,4 % массы взрослых. В четырехмесячном возрасте утки весили 1100 г.
Для выявления особенностей индивидуальной динамики массы крякв был применен метод ежедневных взвешиваний. В опыт были включены как самцы, так и самки. Эмпирическая кривая роста характеризуется крайне резкими колебаниями, имеющимииной вид, чем у кривых, построенных по усредненным показателям более редких взвешиваний (рис. 2, 3). На каждой кривой можно выделить три основных элемента: подъем массы, стабилизация и падение. Сочетание этих элементов характеризует рост каждой птицы, который определяется, помимо генетических факторов, главным образом возрастом птицы и особенностями ее питания.
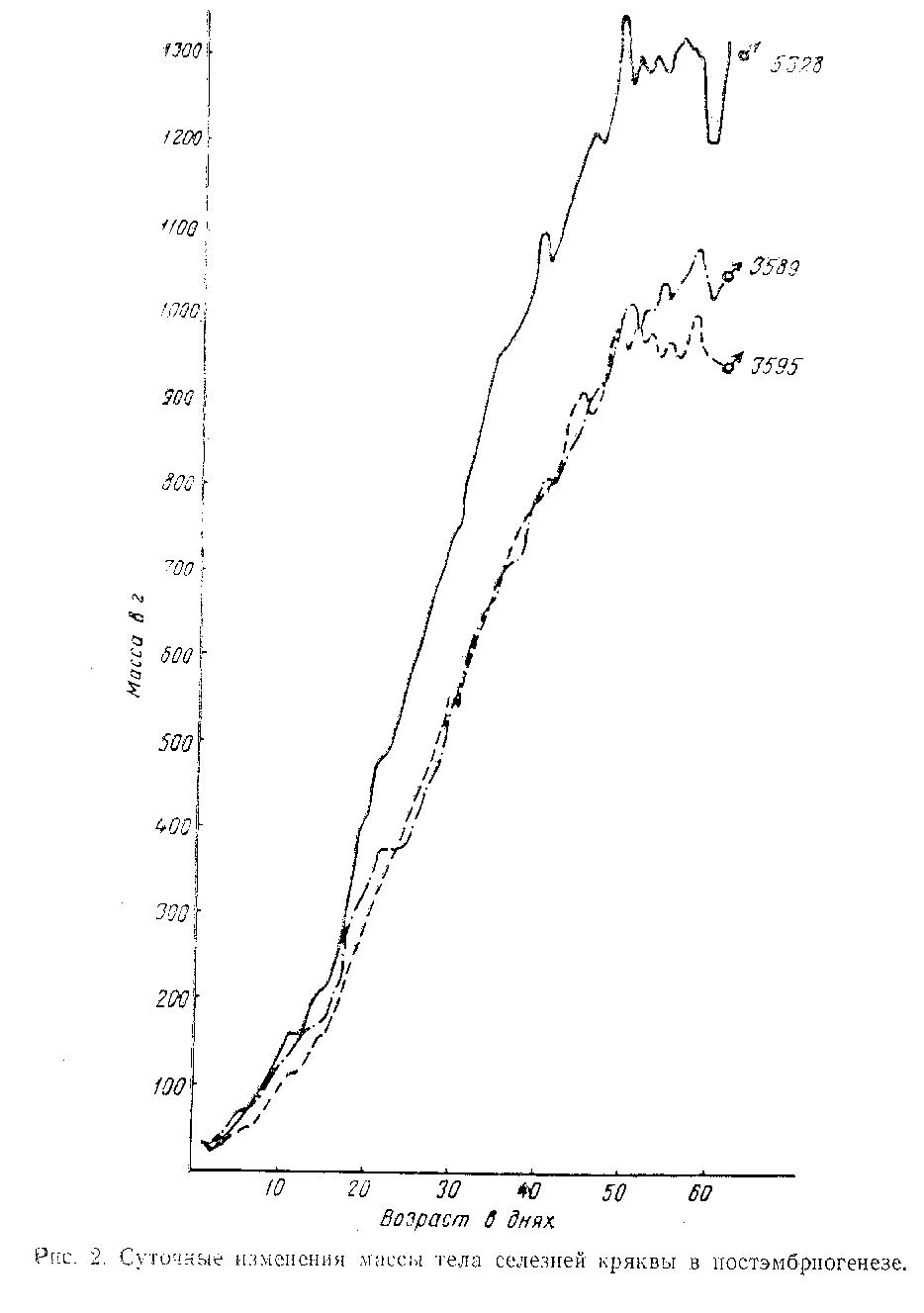

Как видно из кривых роста, общая длина отрезков с подъемом кривой значительно превосходит общую протяженность отрезков приостановки и падения веса. Первые занимают по осиабсцисс несколько дней, приостановка увеличения массы 1-2 дня и падение массы 1-2, иногда 2-3 дня. Кривые массы нa большем своем протяжении представляют собой ломаную линию, что является характерной особенностью графиков, построенных по данным ежедневных взвешиваний. Однако при всем разнообразии в деталях, обнаруживается одна общая закономерность: при нормальном росте крякв резкие падения кривой не опускаются ниже уровня предыдущего падения. Кроме того, острые подъемы кривой почти не выступают выше общего хода кривой и находятся в полном соответствии с предыдущими пиками подъема.
Проведенные исследования характера роста в постэмбриогенезе таких частей тела, как голова, туловище и конечности крякв, показали, что время их роста, так же как иувеличение массы тела, продолжается до определенного возраста птиц и разделяется на несколько этапов, отличающихся разной интенсивностью роста.
Увеличение размеров головы утят по мере их взросления показано в табл. 1.
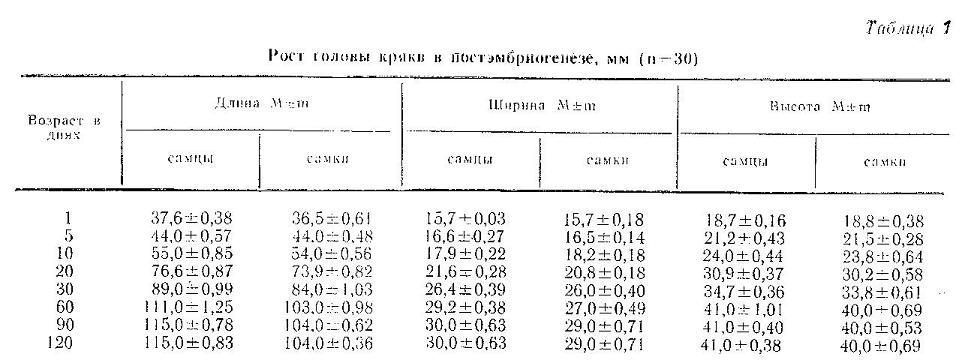
Наиболее интенсивно голова росла вначале постэмбриогенеза. Так, в первые 5 дней относительный среднесуточный прирост головы в длину составлял более 3 %; в следующие 5 дней он достигал максимальной величины более 4 %, далее до 30 дней удерживался на уровне 2 %, после чего снизился и был менее 1 %.
В ширину и высоту голова росла в первом месяце с одинаковой скоростью – 2 % в день, а далее темп роста снижался до полного прекращения к 90-му дню. Кривые их динамики идентичны.
У только что вылупившихся утят пропорции головы отличались от взрослых. Так, длина головы составляла всего 33 % ее длины у взрослых птиц, а ширина и высота - 50 %. У месячных утят длина головы достигала 72-80 %, ширина - около 90 % и высота - более 85 % размеров взрослых. У двухмесячных утят голова по форме уже неотличима от взрослых птиц (длина составляла 99 %, ширина 97 %, а высота уже 100 %), Рост головы в длину и ширину завершался к 90-му дню. У 60-дневных селезней длина головы достоверно отличалась (Р=0,001) от длины головы самок. Остальные показатели не подвержены половому диморфизму.
Рост клюва в длину и ширину у уток представлен в табл. 2. Для выяснения, который из промеров наилучшим образом характеризует развитие клюва в длину, его измеряли в трех разных направлениях (см. примечания к табл. 2). Полученные данные показывают, что наибольшая скорость прироста клюва как в длину, так и в ширину наблюдалась на 10-й день. При этом темп роста клюва в длину по коньку был максимальным (7,1 %) по сравнению с длиной клюва от угла рта (5,9 %) и от ноздри (5,7 %) и, особенно, ростом клюва в ширину (4,3 %). Клюв в длину у селезней рос интенсивнее, чем у уток. С 10 по 30-й день прирост клюва как в длину, так и в ширину снизился, но был еще высоким, а после 30 дней постепенно снижался до полного прекращения к 90-дневному возрасту. Клюв в длину на всем протяжении постэмбриогенеза рос интенсивнее, чем в ширину, и форма клюва по мере взросления утят становилась все более вытянутой.
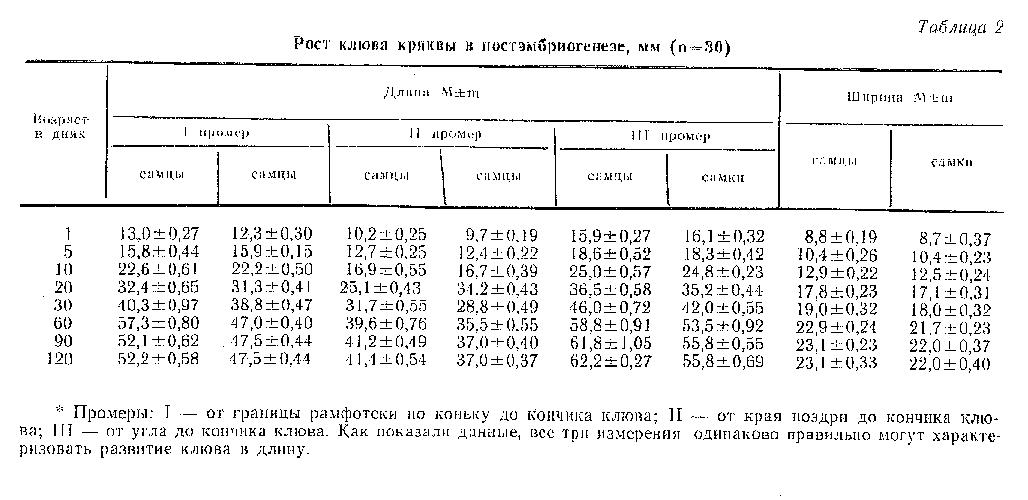
Степень увеличения клюва в длину (по всем трем показателям длины) за период эмбриогенеза составляла 25 %, а в ширину - около 40 %. За первые 30 дней, когда наблюдали высокий темп роста клюва во всех направлениях, длина его достигла уже более 70 %, а ширина более 80 % окончательных размеров взрослых птиц. У месячных селезней длина клюва (по всем трем измерениям) достоверно (Р=0,001) отличалась от длины клюва уток. Различий в ширине клюва между ними не было.
Для выяснения возрастных изменений пропорций туловища крякв изучали рост туловища как в длину, так и в ширину. Полученные данные представлены в таблице 3 и 4. Из приведенных материалов видно, что наиболее интенсивно туловище росло в начале постэмбриогенеза. Так, скорость роста его в длину у селезней и уток в первые 5 дней составляла более 3 %, а в последующие 5 дней была максимальной за весь период постэмбриогенеза - 5,6 %. С 10 по 20-й день темп роста снизился до 2,7 %, а к 30-му дню увеличился у селезней до 3,7 % и уток - до 3,1 %. После 30 дней прирост туловищав длину снижается и к 90-му дню прекращается совсем.
Таблица 3
Рост туловища крякв в постэмбриогенезе, мм (n=30)
|
Возраст в днях |
Длина туловища, M±m |
Ширина груди, M±m |
||
|
Самцы |
Самки |
Самцы |
Самки |
|
|
1 |
46,0±0,64 |
46,0±1,98 |
12,0±0,41 |
12,4±0,42 |
|
5 |
55,0±1,27 |
58,0±1,03 |
14,8±0,65 |
15,0±0,66 |
|
10 |
73,0±1,89 |
77,0±1,04 |
22,0±0,56 |
21,0±0,49 |
|
20 |
97,0±2,10 |
101,0±1,06 |
42,8±1,81 |
40,0±1,85 |
|
30 |
141,0±1,10 |
138,0±3,19 |
43,0±1,85 |
45,0±2,18 |
|
60 |
233,0±5,22 |
224,0±4,09 |
77,0±1,46 |
73,0±1,44 |
|
90 |
245,0±2,89 |
230,0±2,82 |
80,0±0,94 |
75,0±1,16 |
|
120 |
245,0±2,74 |
230,0±2,82 |
80,0±0,94 |
75,0±1,16 |
Таблица 4
Рост туловища в ширину, мм (n=30)
|
Возраст в днях |
Обхват груди, M±m |
Ширина таза, M±m |
||
|
Самцы |
Самки |
Самцы |
Самки |
|
|
1 |
69,0±0,38 |
69,0±1,43 |
14,2±0,42 |
14,8±0,11 |
|
5 |
78,0±1,43 |
78,0±1,49 |
17,0 ±0,41 |
18,4±0,55 |
|
10 |
94,0±2,27 |
98,0±1,43 |
25,0±0,63 |
26,0±0,82 |
|
20 |
180,0±4,68 |
178,0±3,05 |
35,2±0,77 |
35,7±1,00 |
|
30 |
183,0±3,70 |
185,0±3,73 |
44,1±1,02 |
44,8±1,34 |
|
60 |
284,0±5,70 |
278,0±3,36 |
58,1±1,19 |
55,5±1,46 |
|
90 |
292,0±3,16 |
284,0±3,57 |
58,8±1,23 |
56,5±1,72 |
|
120 |
304,0±5,07 |
289,0±1,68 |
58,8±1,23 |
56,5±1,71 |
В первые 20 дней постэмбриогенеза туловище в ширину (в области груди) росло интенсивнее, чем в длину. Так, на 5-й день темп роста ширины груди был более 4 %, а максимальный прирост ширины груди,так же как и длины туловища, наблюдался на 10-й день: у селезней более 7 % и у уток более 6 %. Далее до 20 дней темп роста груди удерживался на высоком уровне (более 6 %), после чего к 30-му дню быстро снижался и с 30 по 60-й день вновь увеличился почти до 2 %. После 60 дней темп роста туловища в ширину снижался и в возрасте 90 дней был равен нулю.
У новорожденных утят длина туловища составляла 19 %, а ширина груди 16 % окончательных размеров взрослых птиц. Поскольку рост туловища в ширину шел интенсивнее, чем вдлину, у месячных птиц ширина груди составляла 54-60 %, а длина туловища около 50 % его длины у взрослых птиц. К 2-месячному возрасту темп роста этих частей тела выравнивался и ширина груди составляла 96 %, а длина туловища 95 % окончательных размеров. У 90-дневных селезней длина туловища и ширина груди достоверно (Р=0,003) отличались от длины туловища и ширины груди самок.
Развитие грудного пояса и изменение с возрастом формы передней части туловища характеризуются не только шириной груди, но и обхватом туловища или груди. У новорожденных утят по сравнению со взрослыми кряквами обхват туловища относительно больше, чем ширина груди и длина туловища, и составляет 24 %. В первые 20 дней жизни утят обхват груди за лопатками увеличивался быстро, достигая максимума к 20 дням (6,3 % у селезней и 5,8 % у уток). К 30-му дню темп роста обхвата груди резко снижался, далее до 60 дней происходил небольшой подъем до 1,5 %, после чего шло постепенное снижение темпа роста до полного прекращения к 120-дневному возрасту. У месячных утят обхват туловища составлял у селезней 60 % и уток 64 % обхвата туловища взрослых птиц, в то время как длина туловища 50 %, а ширинагруди 54-60 %. У 60-дневных утят обхват туловища, также как длина туловища и ширина груди, достигал 96 %, а у 90-дневных утят – 96 % у селезней и 98 % у уток. Четырехмесячные утята имели обхват туловища взрослых птиц. Более продолжительный рост обхвата туловища по сравнению с длиной туловища, очевидно, связан с ростом мускулатуры и уплотнением перьевого покрова.
Увеличение с возрастом таза утят в ширину шло медленнее, чем ширины груди, но быстрее, чем длины туловища. В первые 5 дней жизни темп роста ширины таза был равен темпу роста длины туловища, но ниже темпа роста ширины груди. К 10 дням относительные среднесуточные приросты ширины таза достигали максимума, к 20 дням эта величина снижалась наполовину, но по-прежнему была выше, чем для длины туловища, но ниже, чем для ширины груди. До 30 дней темп роста еще превышал 2 %, а затем постепенно снижался, ирост завершался к 90 дням. Ширина таза новорожденных утят составляла 24-26 % его размеров у взрослых птиц, причем таз был более развит по сравнению с длиной туловища (19 %) и особенно шириной груди (16 %). К месячному возрасту ширина таза у селезней составляла 75 % и у уток 79 % окончательных размеров; ширина груди у селезней 54 % и у уток 60 %, а длина туловища составляла всего 50 % размеров взрослых птиц. К 60 дням таз достигал размеров взрослых птиц, составлял 99 %, ширина груди к этому времени составляла 96-97 %, а длина туловища – 95 %. Полового диморфизма по ширине таза нами не обнаружено.
Возрастная динамика абсолютной длины крыла и предплечья, их относительных среднесуточных приростов приведены в таблице 5. Длина крыла слагается из длины кисти и растущих маховых перьев.
Таблица 5
Рост передних конечностей уток в постэмбриогенезе, мм (n=30)
|
Возраст в днях |
Крыло, M±m |
Предплечье, M±m |
||
|
Самцы |
Самки |
Самцы |
Самки |
|
|
1 |
23,3±0,58 |
21,5±0,72 |
13,3±0,46 |
13,0±0,46 |
|
5 |
25,1±0,66 |
24,9±0,61 |
15,6±0,90 |
15,1±0,60 |
|
10 |
26,2±0,45 |
26,0±0,49 |
17,5±0,51 |
16,7±0,34 |
|
20 |
31,1±1,18 |
33,8±1,45 |
28,7±1,08 |
28,9±0,70 |
|
30 |
58,7±2,55 |
55,7±2,85 |
52,6±2,52 |
51,7±1,56 |
|
60 |
255,0±3,24 |
242,0±4,35 |
89,7±0,72 |
84,5±1,05 |
|
90 |
282,0±2,25 |
273,0±2,22 |
90,0±0,79 |
85,5±0,85 |
|
120 |
282,0±2,25 |
273,0±2,22 |
90,6±1,00 |
85,8±0,96 |
Темп роста крыла и предплечья па всем протяжении постэмбриогенеза неодинаков. В первые пять дней после вывода относительные среднесуточные приросты предплечья составляли 3 %, а крыла - всего лишь 2 %, в следующие пять дней приросты предплечья снизились до 2 % и крыла до 1 %, далее до 30 дней темп роста быстро увеличивался и у 30-дневных птиц достигал максимальной величины – 6 %. После месячного возраста темп роста предплечья снижался быстрее, чем крыла, и к 90-дневному возрасту приближался к нулю.
Передняя конечность по сравнению со всеми изученными структурами у новорожденных утят менее всего развита: предплечье составляло 15 %, а крыло лишь 8 % конечных размеров. Предплечье месячных утят достигало 60 %, а крыло всего лишь 20 %. Из данных видно, что сама передняя конечность (предплечье) имела темп роста в 2,0 раза выше, чем крыло. Крыло оказалось менее развитым потому, что его длина зависела от роста не только кистевой части скелета передней конечности, но и от роста маховых перьев, которые к этому времени еще недостаточно выросли. С 30-го по 60-йдень происходит бурный рост перьевого покрова, и у 2-месячных птиц длина крыла составляла уже 90 % длины его у взрослых птиц. Предплечье двухмесячной птицы почти достигло окончательных размеров (99 %). Длина предплечья 60-дневных селезней статистически достоверно (Р=0,001) отличалась от длины предплечья уток. Полного развития крыло достигло к 3-месячному возрасту. Достоверных различий в длине крыльев у 120-дневных селезней и уток нами не обнаружено.
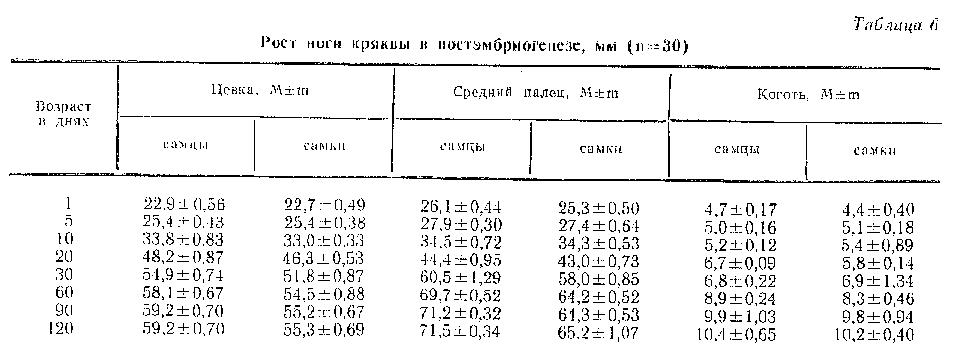
О возрастных изменениях длины цевки, среднего пальца и когтя крякв можно судить по данным таблицы 6. Результаты измерений показывают, что ноги у утят наиболее интенсивно росли в первые 10 дней, причем максимальный прирост цевки и среднего пальца наблюдался на 5-й день. Темп роста цевки был выше темпа роста среднего пальца. После 10 дней темп роста конечности снижался и у 20-дневных птиц составлял: цевки - 3,5 %, среднего пальца - 2,2 %. После 20 дней темп роста цевки постепенно снижался до полного прекращения к 90-му дню. Темп роста среднего пальца и когтя к 30-му дню несколько повышался (у среднего пальца до 3 % и у когтя до 2 %), после чего начинал постепенно снижаться. Все части задней конечности крякв к моменту вывода были наиболее развитыми в сравнении с другими частями тела и составляли около 40 % конечных размеров у взрослых птиц. У месячных утят ноги были самым развитым органом из всех изученных нами частей тела, так как цевка составляла более 90 %, а средний палец около 90 % размеров взрослых птиц. Крыло в этом возрасте составляло всего лишь 20 % окончательных размеров. Это объясняется тем, что утки, как и все водоплавающие, почти сразу после вылупления при помощи ног могут легко перемещаться по суше и, особенно, плавать в воде, а крылья у них начинают функционировать значительно позже. У 2-месячных утят крыло составляло 90 % размеров крыла взрослых птиц, а цевка и средний палец уже почти закончили свой рост и составляли 98 % размеров взрослых птиц. У 60-дневных птиц длина цевки и среднего пальца селезней достоверно (Р=0,004 и Р=0,001) отличались от длины цевки и среднего пальца самок.
На основании полученных данных можно сделать следующие выводы.
1. Нарастание массы крякв в постэмбриогенезе продолжалось до 3-месячного возраста.
2. Индивидуальные изменения абсолютной массы тела крякв в постэмбриогенезе разделяются на три этапа: первый - подъем массы тела (длится от 3 до 7 дней), второй - приостановка нарастания массы тела (длится 1-2 дня) и третий - падение массы тела (длится 1-3 дня).
3. Скорость весового роста крякв изменялась обратно пропорционально возрасту утят и имела волнообразный характер, состоящий из трех пиков на 10, 20, 30-й дни.
4. В 2-месячном возрасте масса селезней статистически достоверно отличалась от массы тела уток.
5. Новорожденные утята имели высокие ноги, относительнотбольшую и широкую голову, сравнительно широкий клюв, короткое тело и короткие крылья.
6. Рост частей тела утят, так же как и нарастание массы тела, в постэмбриогенезе продолжался в основном до 3-месячного возраста.
7. Скорость линейного роста, так же как и весового, изменялась обратно пропорционально возрасту утят. Наибольшая скорость наблюдалась в начале постэмбриогенеза (от 10-20 дня), затем она начинала постепенно снижаться до полного прекращения. Одни части тела характеризовались максимальным приростом к 10-дневному возрасту (длина головы, длина и ширина клюва, длина и ширина туловища, длина цевки и среднего пальца). Прирост других достигал максимума к 20-дневному возрасту (длина крыла и предплечья).
8. Половой диморфизмпроявился:
а) в месячном возрасте - по длине клюва;
б) в двухмесячном возрасте - по длине головы, предплечья, цевки и среднего пальца;
в) в трехмесячном возрасте - по длине и ширине туловища.