
Перерва В.И., Киселёва Е.Г. Деформация репродуктивных циклов при разведении зубра (Bison bonasus L.) в неволе // Дичефермы и зоопитомники. Сборник научных трудов ЦНИЛ Главохоты РСФСР. М., 1991. С. 165-179.
Одним из важных аспектов проблемы сохранения генофонда зубра является восстановление естественных видовых репродуктивных циклов у современных животных. Существенные изменения физиологии размножения вида произошли в связи с тем, что многие поколения этих зверей подвергались интенсивному воздействию условий неволи. Они проявились в первую очередь в растянутости сроков гона и отела. Аборигенным зубрам Беловежской пущи и Кавказа в прошлом веке был присущ короткий период половой активности (гон, яр), длившийся 2-3 недели - с конца августа по конец сентября (Brincken, 1826; Усов, 1888). Но уже на исходе XIX столетия появились сведения об удлинении этих сроков. Г. Карпов (1903) отмечал, что в то время гон у зубров протекал уже на протяжении 1,5 месяцев - с июля по сентябрь. Н.М. Кулагин (1919) называет еще больший интервал - с июня по октябрь, т.е. гон зубров удлинился почти до 3 месяцев.
В последующие десятилетия у животных в зверинцах, зоосадах, питомниках период гона и отелов деформировался так, что исчезла ярко выраженная сезонность, видоспецифичная для всех диких зверей (Jaczewski, 1958). Судя по замечаниям К. Вроблевского (Wroblewski, 1927) о том, что в Беловежской пуще в начале XX столетия позднелетние и зимние отелы одинаково часто встречались как у вольных, так и находившихся в зверинцах животных, налицо единый процесс разрушения естественного репродуктивного цикла у последних аборигенных зубров.
Исследования в указанном направлении важны по ряду причин. Не оправдываются надежды на быстрое и полное восстановление естественной циклики размножения зубров после их выпуска в природу. На это указывают результаты первых десятилетий вольного обитания животных в Беловежской пуще как белорусской ее части (Корочкина, 1971; Буневич, Кочко, 1988), так и польский (Пуцек, 1986). К вольным зубрам пока еще не возвратился характерный для аборигенных популяций прошлого века исключительно майский пик отелов. И сегодня в ряде вольных стад одним из серьезных последствий расшатанности циклов размножения является рождение телят в позднеосеннее время, что ставит их в трудные условия зимовки. Вскрыв действительные причины, вызвавшие такую трансформацию физиологии современных зубров, можно попытаться вернуть им естественный ритм размножения путем конкретных практических мер. Последнее обстоятельство крайне важно для успешного завершения программы восстановления вида в природе.
В основу представленного анализа лег опыт разведения чистокровных беловежских и кавказско-беловежских зубров, а также их гибридов с бизоном и крупным рогатым скотом в трех специализированных питомниках СССР: в Приокско-Террасном (год основания - 1948) и Окском (1969) заповедниках, а также в госохотхозяйстве «Науяместис» Литовской ССР (1972 г.). В этих резерватах до 1988 г. зарегистрировано следующее число актов рождения зубрят: в Приокско-Террасном заповеднике в Центральном зубровом питомнике (ЦЗП) - 395 чистокровных зубров обоих форм и 132 гибрида и бизона, в Окском заповеднике (ОГЗ) - 256 кавказско-беловежских, а в ГОХ «Науяместис» - 74 беловежских зубра. Особенностью разведения в питомниках является строгий учет всех случаев рождения, мертворождения, гибели с точной регистрацией дат этих событий, что находит отражение в издаваемой в Варшаве European Bison Pedigree Book. Это позволило провести статистическую обработку материалов, имеющихся в племенных книгах. Основное внимание обращено на сроки отелов и гона. Для определения сроков гона мы пользовались информацией, характеризующей половую активность зубров обоих полов (число копуляций в течение каждого месяца): для ОГЗ такие данные имеются с 1961 г., а для ЦЗП - с 1975 по 1978 г. В ряде случаев гон оценивался по результатам рождения телят на основе восстановления сроков оплодотворения, когда от даты отела вычиталось 262 дня - средняя продолжительность беременности зубров в ЦЗП (Заболоцкий, 1957). Для выявления связи этих репродуктивных параметров с соответствующими факторами использовалась как графическая форма обработки материала, так и статистическая - рассчитывались коэффициент корреляции - r и коэффициент вариации - cv. Определялась зависимость сроков гона от погодных условий, в частности, от температуры воздуха (использованы величины среднемесячной температуры). Метеоданные брались со стационарных метеостанций, расположенных в Приокско-Террасном заповеднике в 300 м от границ питомника, а в Окском заповеднике - в 1 км.
Для зубров в дикой природе, как и для всех копытных (Rutberg, 1984), естественным является короткий период прохождения гона и связанных с ним последующих отелов. Первой реакцией на изменение условий жизни зубров, оказавшихся в неволе, была расшатанность репродуктивных циклов. Наиболее показательны в этом плане сроки отела зубриц. У содержащихся в питомниках зубров в настоящее время сформировались репродуктивные циклы, растянутые почти на весь календарный год. На рис. 1 представлены кривые, демонстрирующие сезонность отелов у зубров, живущих и живших в различных питомниках СССР, а на рис. 2 - динамика изменения сезонности гона с первых лет существования зубрового питомника в Окском заповеднике. Во всех бывших и ныне функционирующих резерватах СССР отелы зубров проходили почти во все месяцы года, исключая, может быть, лишь февраль. Однако наиболее часто (56,1% всех зарегистрированных случаев) телята рождались в апреле-июне. Почти такая же цифра (55,8 %) для тех же сроков была получена Jaczewski (1958) для зубров в питомниках Европы в период до 1953 г. Если следовать выводам автора, считавшего естественным для вида отел в мае-июне, то в питомниках во все периоды их существования лишь немногим более половины самок телились в нормальные сроки. Остальные случаи следует рассматривать как результат расшатанности репродуктивного цикла зубров.
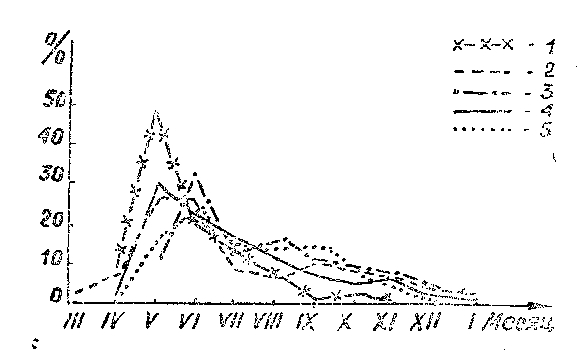
Рис. 1. Сезонность отелов зубров в некоторых питомниках СССР: 1 - Кавказский заповедник (1940-1964 гг.) по С.Г. Калугину, 1968; 2 - Беловежская пуща (1946-1965 гг.) по Л.П. Корочкиной, 1971; 3 - Охотхозяйство «Науяместис» (1970-1983 гг.); 4 - Приокско-Террасный заповедник (1948-1988 гг.); 5 – Окский заповедник (1969-1988 гг.).
Темпы изменения циклики зубров значительны, судя по гистограмме, показывающей половую активность животных в ОГЗ (рис. 2). Уже на восьмой год работы питомника гон затягивался до середины зимы (январь), а на 17 год это явление стало регулярным и, более того, распространилось на февраль-март.

В данной статье обсуждается влияние на репродуктивный цикл зубров некоторых технических приемов, употребляемых в зуброводстве. Основной механизм изменения циклики заключается в том, что самки не могут оплодотвориться в нормальные для вида сроки - август-сентябрь. Так как у зубриц в течение года может наступать несколько течек (эструс), то пропуск первой из них ведет к затягиванию гона на более позднее время осени. Самки, у которых оплодотворение происходит в осенние или тем более в зимние месяцы, попадают в неблагоприятные погодные условия, что в свою очередь тормозит нормальное половое развитие. В результате позднее оплодотворение определяет поздний отел, а поздний отел в свою очередь еще дальше отодвигает дату следующего оплодотворения.
Э. Ячевски (Jazewski, 1958) считает, что у зубров интервал между отелом и последующим оплодотворением должен быть не менее 100 дней, т.е. немногим более 3 месяцев. Но в зубровых питомниках период между отелом и последующим оплодотворением сокращается в среднем до 40 дней, а в исключительных случаях и до 22 дней. Такое сокращение обусловлено тем, что в неволе практикуется своеобразная стимуляция высокой плодовитости племенных животных. Условиями для этого являются, с одной стороны, постоянное присутствие быка в одном загоне с самками, а с другой, отъем телят в возрасте 10 месяцев от матерей, у которых еще не закончилась лактация. Конечно же, за тот короткий срок, который дается зубрицам для отдыха после отела, их организм не успевает полностью восстановиться к повторной беременности. Вследствие этого в первую течку после запоздалого отела оплодотворение скорее всего не происходит. Оно произойдет во вторую или даже в третью течку, т.е. с опозданием на 1-2 месяца относительно нормальных для вида сроков. Начинается накопление отсрочек как оплодотворения, так и отелов.
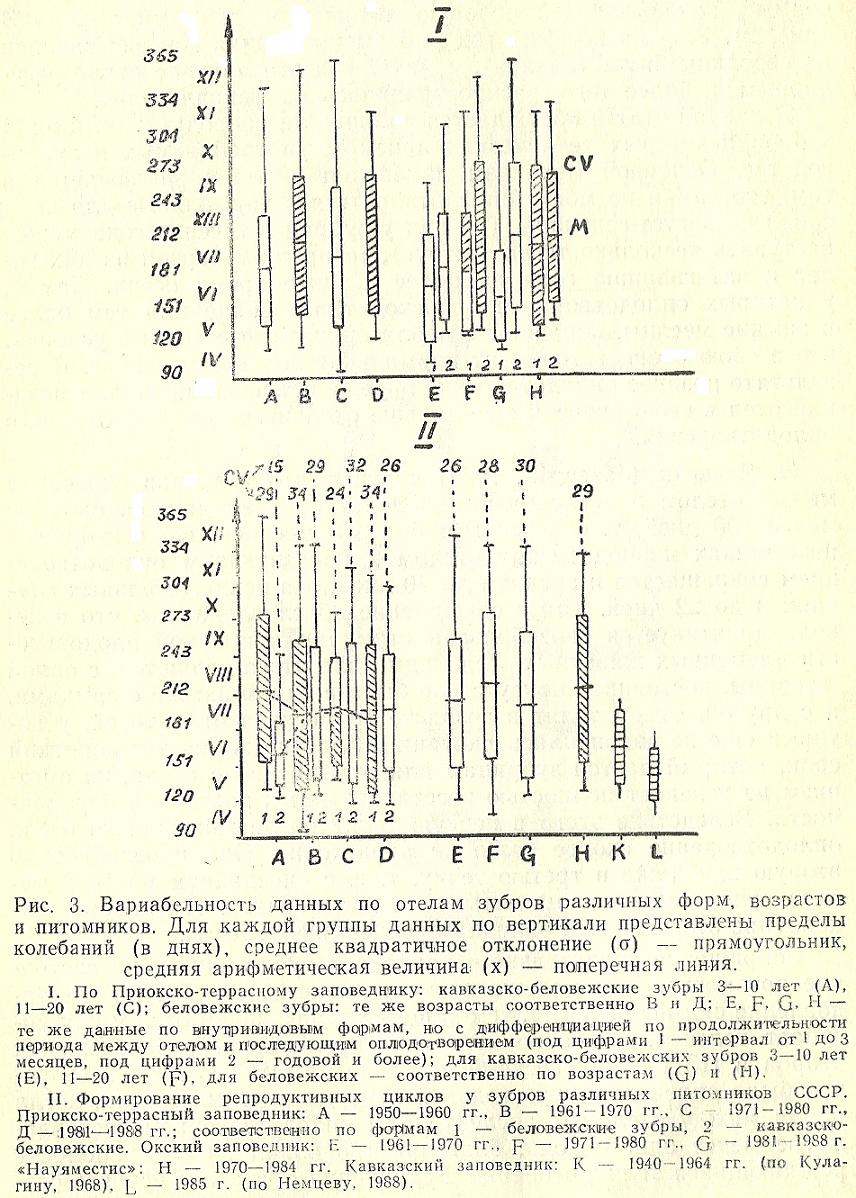
Сокращение предотельного периода пагубно влияет на состояние потомства. Чем меньше времени остается самкам на восстановление сил после отела, тем чаще в последующий отел у них погибает теленок. Особенно четко это проявляется у беловежских зубров в ЦЗП и кавказско-беловежских в ОГЗ. С другой стороны, было определено, что годовой перерыв в размножении, когда время между отелом и последующим оплодотворением составляло не менее одного года, благоприятно влиял на выживаемость рождающихся телят, особенно в сравнении с успехом первого отела зубриц в тех же питомниках. После такого перерыва зубрицы обычно телились в апреле-июне (65-70 % всех отелов в течение года), т.е. к ним возвращался нормальный репродуктивный цикл. Примечательно, но и беловежские и кавказско-беловежские самки одинаково реагировали на такое прохолостание. Даже статистическая обработка данных материалов показывает тенденцию к такому сдвигу, хотя достоверность различий не подтверждена по t-критерию (рис. 3). Однако если сравнивать средние арифметические дат рождения телят молодыми зубрицами 3-10-летнего возраста (Е2 для кавказско-беловежских и G2 для беловежских) в периоды ежегодных отелов с датами рождения телят самками тех же возрастов, но после годового перерыва (El и G1, соответственно по внутривидовым формам), то первые оплодотворяются и телятся на месяц раньше вторых.
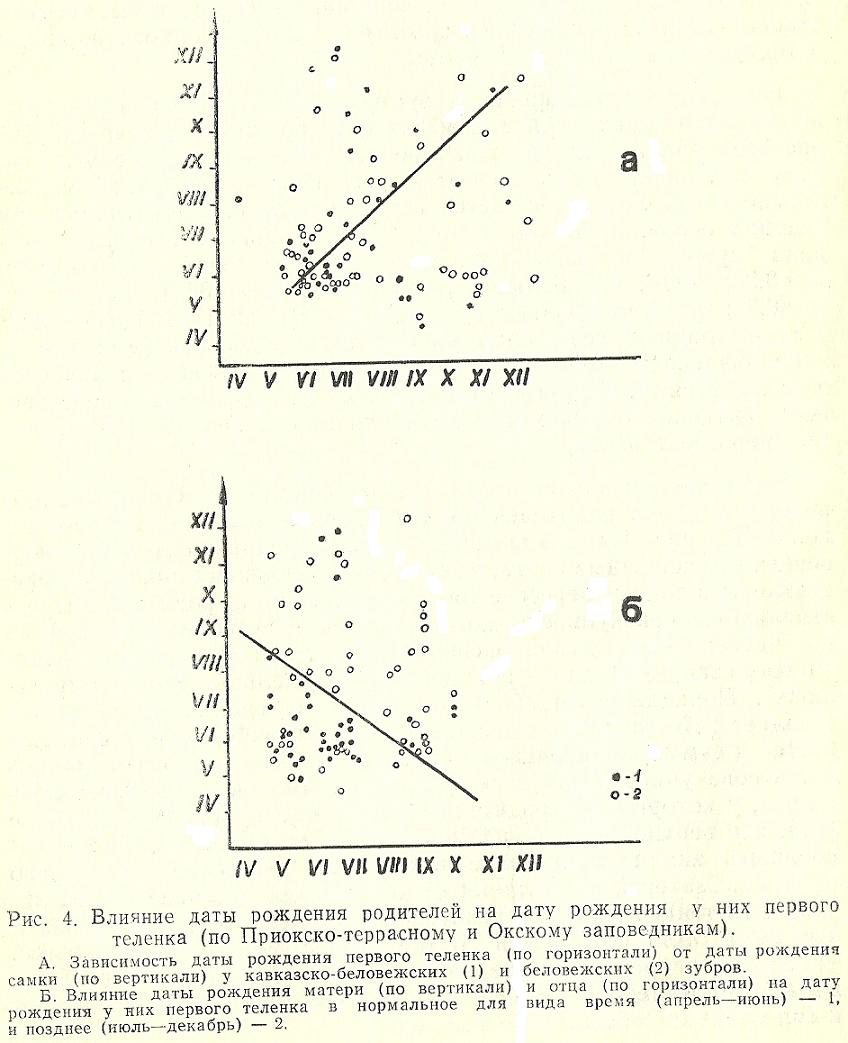
На циклику размножения зубров влияет подбор племенных животных по таким признакам как дата рождения и возраст. Для расчетов брались даты только первого отела каждой самки, потому что, на наш взгляд, в этот момент и ни в какой другой из последующих отелов проявляется естественный репродуктивный потенциал особи. Была определена прямая зависимость даты рождения зубрят от сроков рождения их родителей как матери (r=0,93; t=16, что больше tst=3,37 при Р=0,001), так и отца (r =0,94; t=16, что больше tst=3,37 при Р=0,001). В ЦЗП и ОГЗ у самок, родившихся в нормальные для вида месяцы (май, июнь), в подавляющем большинстве случаев первый теленок появляется в те же месяцы. С другой стороны, более поздние сроки собственного рождения самок могут быть причиной позднего рождения у них первого теленка.
Была сделана попытка оценить одновременное влияние сроков рождения обоих родителей на дату появления у них первого теленка. На рис. 4 «Б» нормально и позднородившиеся телята-первенцы, обозначенные соответствующими условными значками, расставлены в точках пересечения дат рождении их родителей. Представленную совокупность можно разделить условной прямой на две части: выше прямой располагаются позднорожденные телята (июль-декабрь), ниже - рожденные в нормальные сроки (апрель-июнь). Попадание «ошибочных» особей за границу раздела составляет 25 % (84:21 - отношение суммы особей обеих совокупностей к сумме «ошибочных» особей, попавших в противоположную часть совокупности). В нижней части совокупности разместились телята, у которых оба родителя были рождены весной и в начале лета, а в верхней - в более позднее время. Однако дифференцированный анализ представленных частей совокупности отдельно по линии матерей и по линии отцов не дали достоверных различий, хотя тенденция к рождению поздних телят родителями, которые сами были рождены в поздние сроки, все-таки сохраняется. Такое противоречие с материалами на рис. 4 «А» обусловлено, видимо, проявлением компенсирующего влияния родителей с разными сроками собственного рождения.
Сроки размножения зубров в питомниках удлиняются и при участии в гоне старых особей. Животные старше 10 лет оплодотворяются позднее молодых (рис. 3 «1» А-В, С-Д). Тенденция может быть обусловлена тем, что у старых особей течка задерживается до октября-ноября и потому позднеосенние похолодания отодвигают сроки их оплодотворения еще далее на неблагоприятное зимнее время.
Существенным фактором изменения репродуктивной циклики зубров являются погодные условия местообитания животных, даже если это относится к питомникам. В данном сообщении представлен материал, демонстрирующий связь размножения с температурой воздуха. Определена основная закономерность такой взаимосвязи: с 1 октября по 31 декабря с понижением температуры происходит снижение половой активности зубров и, наоборот, с повышением температуры гон возобновляется (r =0,81; t=9,95, что больше tst при Р=0,001).
В Подмосковье и Рязанской области при сильных похолоданиях, происходящих либо в октябре, либо в ноябре, резко снижается половая активность зубров обоих полов. И чем сильнее понижалась температура, тем меньше регистрировалось число актов ухаживания самцов за самками (1975 и 1976 гг. в обоих питомниках). Иногда гон совершенно прекращался, однако, в последующие недели половая активность возобновлялась и проходила порой так же интенсивно, как в августе-сентябре. На такой всплеск активности влияет и некоторое улучшение погоды, например, потепление, если оно бывает, и адаптация самих зверей к суровому времени года. Из этого следует, что похолодание прерывает гон зубров, заставляя тех самок, которые в теплое время осени не успели -оплодотвориться, вновь объединиться с быками, но в более позднее время. В питомниках это возможно, ибо быки постоянно находятся со стадом зубриц, а на воле такая связь затруднена из-за привычки самцов уединяться сразу же по окончании гона.
Взаимосвязь циклики размножения с погодой в местах обитания зубров - это лишь составная часть более общего процесса влияния физико-географических условий на физиологию животных. Под физико-географическими условиями мы понимаем весь комплекс абиотических факторов, к которым должны адаптироваться зубры, насильно переселяемые в новые места существования. В результате у них формируются специфические для данной местности репродуктивные циклы (рис. 1). В южном питомнике, каким был до 1965 г. питомник в Кавказском заповеднике, зубры телились, в большинстве случаев, в июне (рис. 3 «II» К.). Однако процесс формирования оптимальных сроков гона и отела к тому времени еще не завершился и в последующие два десятилетия происходила дальнейшая адаптация, но уже в условиях полной свободы (Немцев, 1988). В результате к середине 1980-х годов определялся четкий майский пик отелов (рис. 3 «II» L), который, видимо и является естественным для данного региона.
В западных питомниках (в белорусской Беловежской пуще и в литовском охотхозяйстве «Науяместис») отелы проходили одинаково часто как в мае, так и в июне, но немало их было и в июле-августе. В целом, для самого северного питомника СССР («Науяместис») оказалось, что отелы наблюдались в среднем в наиболее поздние сроки (рис. 3 «II» Н), чем в других местах.
Не менее растянутым оказался период размножения у зубров в восточных питомниках - в ЦЗП и ОГЗ, особенно в последнем, где одинаково часто рождались телята на протяжении 4 месяцев - с мая по сентябрь (рис. 1).
На формирование оптимальных для региона сроков гона и отела необходимо достаточно большое время. Сравнение графиков прохождения отелов у различных форм зубров (беловежских и кавказско-беловежских) из ЦЗП и ОГЗ (рис. 3 «II») показывает, что независимо от различий в изначальном репродуктивном потенциале привезенных из Польши животных, в последующем происходило формирование единого для соответствующей местности ритма размножения. В ПТЗ в 1950-1960-е годы кавказско-беловежские зубры телились в большинстве случаев в мае, тогда как беловежские - в июле. Но в третье и, особенно, в четвертое десятилетия работы питомника произошло выравнивание репродуктивных циклов обеих форм и кривые отелов стали иметь одинаковую конфигурацию. Аналогичным образом происходило приспособление кавказско-беловежских зубров к условиям Окского заповедника.
Нужно признать, что процессы трансформации различных аспектов биологии зубра мало исследованы, ибо до последнего, времени не придавалось серьезного значения возможным последствиям для вида длительного разведения в неволе. Все работы с данным объектом были пронизаны настоятельной необходимостью сохранения вида всеми доступными способами. В настоящее время, когда зубр считается восстановленным видом и ему не грозит исчезновение по причине малочисленности, актуальным становится сохранение естественных видоспецифических черт его биологии. Все чаще появляются факты, свидетельствующие о наличии у современных животных значительных отклонений тех или иных признаков от естественных, которые были присущи истребленному предку. Возвращение животных в современную среду обитания не привело к автоматическому восстановлению деформированных признаков, что усложняет решение проблемы восстановления зубра в диком состоянии. Не исключено, что в будущем нужны будут конкретные работы по искусственному стимулированию процессов регенерации естественных видовых особенностей.
Такой взгляд на существо проблемы не разделялся в прошлом и не всеми признается в настоящее время. Это и понятно - для того, чтобы уловить столь тонкие процессы, требуется накопление большого фактического материала и иной уровень анализа. Еще в самом начале работ по восстановлению зубров (1930-1940 гг.) появились первые признаки доместикации животных, разводимых в неволе. Это было общеизвестно, однако этим фактам противопоставлялось априорное, но категорическое суждение об обратимости процессов деформации экологических характеристик вида. Эти две противоположные точки зрения постоянно конкурировали друг с другом. Так, в одной из своих первых работ М.А. Заблоцкий (1949), анализируя накопленный к тому времени опыт разведения зубров в ряде питомников, делает вывод о появлении определенных изменений в ритмике размножения вида. Однако Z. Jaczewski (1959), изучив размножение зубров в резервах Польши, не обнаружил необратимых изменений репродуктивного потенциала вида. По целому ряду признаков (плодовитости, соотношению полов, сроках наступления половозрелости и т.п.) эту точку зрения разделяла и Л.Н. Корочкина (1971) на основе обобщения результатов исследований в СССР. Позднее ряд авторов возвращались к данной теме (Krasinski, Raczynski, 1969; Киселева, 1969; 1973) и обращали внимание на серьезную трансформацию некоторых признаков физиологии размножения вида в питомниках Польши и СССР. К этому следует добавить анализ размножения горных зубров в Кавказском заповеднике (Калугин, 1968) и чистокровных животных в Центральном зубровом питомнике (Перерва, 1981, 1983).
Одним из первых рецидивов более чем столетнего существования вида в неволе явилось разрушение репродуктивного цикла, выработанного в процессе эволюции зубра. У аборигенных популяций Беловежской пущи и Кавказа очень короткий, всего лишь 2-3-недельный, период гона позволял самкам отелиться в сжатые сроки конца весны, а появившемуся потомству быстро окрепнуть в наиболее благоприятное летнее время года. За последнее столетие произошла полная деградация этого важного признака, что было расценено многими исследователями как проявление доместикации зубра.
Как предполагает Z. Jaczewski (1958), однообразная, но регулярная подкормка в качестве пускового механизма доместикации была основным фактором, кардинально изменившим репродуктивный цикл домашних коров. По аналогии с этим процессом влияние круглогодичной подкормки было перенесено и на зубра. К данному фактору некоторые авторы добавляют еще и обеднение кормовой базы как в природе (в Беловежской пуще), так и в загонах питомников, где содержались звери (Заблоцкий, 1949). В какой-то мере такая точка зрения находит подтверждение в том, что кавказские зубры, никогда не имевшие искусственной подкормки, и в начале XX столетия проводили гон в короткие срои-с середины августа по конец сентября (Башкиров, 1940).
Именно с возвратом в леса и переходом на питание естественными кормами исследователи связывали надежды на возвращение зубрам нормальной циклики размножения (Заблоцкий, 1949). Однако процесс регенерации репродуктивного потенциала вида в современных вольных популяциях проходит крайне медленно. В польской части Беловежской пущи по сведениям З. Пуцека (1986) на три месяца (май-июль) приходится 81 % всех отелов свободно живущих зверей, что лишь на 18 % больше, чем аналогичные показатели для зубров, находящихся в питомниках и зоопарках. Еще более растянут период отелов в популяции зубров белорусской части Пущи (Бруневич, Кочко, 1988). Здесь одинаково часто самки телились в течение четырех месяцев (май-август). И только в одном регионе завершился процесс восстановления естественной ритмики размножения - в Кавказском заповеднике (Немцев, 1988). Однако процесс этот был не однозначен. В период загонного содержания горных зубров (1941-1955 гг.) для них был характерен явный майский пик отелов, тогда как в первые годы вольного обитания (1956-1960 гг.) он сдвинулся на июнь (Калугин, 1968). К 1985 г. появление телят в мае вновь стало массовым явлением (Немцев, 1988).
Из выше приведенных сведений нельзя судить о прямой связи ритмики размножения зубров исключительно лишь с трофическим фактором. Если применительно к зубровым питомникам можно признать зависимость явления смещенности сроков размножения от однообразия подкормки и недостатка естественных кормов, то сохранение данной тенденции при полной свободе выбора пищевых ресурсов в естественных лесах Беловежской пущи, да и в первые годы обитания на воле в Кавказском заповеднике, на наш взгляд, демонстрирует более сложную природу данного процесса.
Еще одним мощным фактором, расшатывающим биологию зубра, признается инбридинг. Однако применительно к циклике размножения этому фактору не придавалось существенного значения. Правда, уже в 1950-е годы З. Ячевски (Jaczewski, 1958) обратил внимание на более выраженный осенний сдвиг отелов в высокоинбредном стаде беловежских зубров в питомнике Пшина в сравнении с другими племенными группами. Но в итоге автор категорически исключил возможность трансформации размножения зубров под влиянием инбридинга, отдав предпочтение в этом трофическому фактору. Лишь в последние годы детальный статистический анализ зависимости коэффициента инбридинга от сезона рождения зубрят показал высокую степень их корреляции (Olech, 1987). Однако из указанной работы невозможно определить, что же является первопричиной: то ли, чем выше коэффициент инбридинга, тем позднее рождаются телята, то ли наоборот. В то же время ответ на данный вопрос имеет принципиальное значение. Ведь, судя по результатам исследования В. Олех (Olech, 1987), в последние годы в мировой популяции зубров происходит снижение инбредности. В таком случае следует ожидать и автоматического возвращения репродуктивной циклики зубров в рамки естественных сроков, что, однако, весьма сомнительно, по крайней мере для двух основных питомников СССР (ЦЗП и ОГЗ).
Ни в коей мере не умаляя влияния выше указанных причин (трофика и инбридинг), мы обращаем внимание на некоторые технологические приемы содержания зубров в неволе, на подбор племенных особей, а также на географию размещения племенных центров в качестве факторов, трансформирующих размножение. Недооценка негативного их воздействия на биологию размножения существенно сдерживает процесс регенерации естественных видовых признаков зубра. Искусственная стимуляция ежегодной рождаемости зубрят, для чего в корне меняется социальная структура стад (круглогодично содержатся вместе половозрелые быки и коровы), насильственно прерываемая лактация самок при отъеме 10-месячных телят - все это ведет к удлинению сроков половой активности, пропуску течек в нормальные сроки (август-сентябрь) и в конечном итоге обуславливает рождение слабых телят в неблагоприятное время года. Необходимо категорически отвергнуть тезис о естественности для зубров ежегодных отелов, который лежит в основе указанного технологического приема разведения (Заблоцкий, 1957). Уже в 1950-1960 гг. многие исследования подтверждали, что интервал между отелами у зубриц на 2-3 месяца более года (Krasinski, Raczynski, 1967; Корочкина, 1971). В одном этом уже заключен механизм чередования отелов через год прохолостания, который необходим самкам для нормального выкармливания теленка хотя бы до 1,5 лет. Искусственная стимуляция ежегодной оплодотворенности нарушает естественный ход развития физиологических процессов в организме самок, что проявляется в ряде отклонений, о которых было указано в наших исследованиях.
Имеется обширная литература, анализирующая влияние возраста родителей на состояние потомков, и в первую очередь, на их пол (Заблоцкий, 1957; Калугин, 1958; Jaczewski, 1958; Корочкина, 1968). Однако нас интересовала связь возраста и сезона рождения родителей на сроки появления потомства. Представленный материал достоверно подтверждает, что у зубров, рожденных в нормальные для вида сроки, потомство также рождается в аналогичное время. И наоборот.
Еще одним важным фактором трансформации репродуктивных циклов являются климатические и погодные условия местности, в которой расположен питомник. К сожалению, этот фактор в литературе по зубру почти не анализировался. Лишь М.А. Заблоцкий (1949) вскользь упомянул о наличии различий в сроках отела, а значит и гона, в резерватах различного географического положения, но детального анализа не провел. Ранее мы показали влияние погодных условий на линьку зубров (Перерва, 1982), когда была определена прямая корреляция данного аспекта физиологии зубров с таким абиотическим фактором как температура воздуха. Есть предположение о том, что климат является решающим фактором при формировании оптимальных сроков отела у бизонов (Rutberg, 1984), хотя доводы в пользу такого влияния, приведенные автором, малоубедительны. Наши материалы показывают обратный процесс, а именно, что погода сама по себе и в комплексе с физико-географическими условиями местности, где размещается племенной центр, может оказаться существенной причиной разрушения нормальной циклики размножения зубров. Такое разрушение сопровождается формированием иной циклики половой активности животных, которая, на наш взгляд, является оптимальной для данного региона. В этом заключена суть адаптации зверей к новым условиям жизни.
Проведенный анализ племенной деятельности зубровых питомников в СССР позволяет наметить комплекс практических мер, реализация которых, на наш взгляд, может вернуть зубру нормальную циклику размножения. Для этого нужно вести племенную работу по следующим направлениям:
1. Подбор самок должен учитывать дату их рождения. В племенное стадо нужно включать только зубриц, родившихся в мае-июне.
2. В племенных стадах не должно быть животных старше 15 лет.
3. Самцы должны удаляться из племенных групп на полгода с октября по июнь и допускаться к самкам только на период гона.
4. Проявление поздней беременности у зубриц должно сопровождаться последующим искусственным перерывом в размножении не менее чем на один год.
ЛИТЕРАТУРА
Башкиров И. Кавказский зубр // Кавказский зубр. М., 1940. С. 3-72.
Буневич А.Н., Кочко Ф.П. Динамика численности и структура популяции зубров Беловежской пущи // Популяционные исследования животных в заповедниках. М.: Наука, 1988. С. 96-114.
3аблоцкий М.А. Необходимость изучения особенностей зубра и его восстановление в СССР // Научно-метод. зап-ки. М., 1949. Вып. 13. С. 128-l46.
3аблоцкий М.А. Некоторые биологические особенности зубра и их изменение в условиях загонного содержания // Тр. Приокско-Террасного заповедника. М., 1957. Т. 1. С. 5-65.
Калугин С.Г. Зубры в естественных условиях Кавказского заповедника // Тр. Кавказского заповедника. Майкоп, 1958. Вып. 4.
Калугин С.Г. Восстановление зубра на северо-западном Кавказе // Тр. Кавказского заповедника. М., 1968. Вып. 10. С. 3-94.
Карцев Г.Л. Беловежская пуща. Спб, 1903. 414 с.
Киселев Е.Г. Материалы о размножении и плодовитости зубров в Окском заповеднике // Редкие виды млекопитающих фауны СССР к их охрана. М.: Наука, 1973. С. 123-124.
Корочкина Л.Н. Факторы, влияющие на соотношение пола у зубров // Беловежская пуща. Минск. 1968. Вып. 2. С. 139-147.
Корочкина Л.Н. Показатели размножения зубров в Беловежской пуще // Беловежская пуща. Минск, 1971. Вып. 4. С. 176-184.
Кулагин Н.М. Зубры Беловежской пущи: Сб. Московского научного института. М., 1919. 166 с.
Немцев А.С. Особенности биологии, охрана и пути использования популяции горных зубров северо-западного Кавказа: Автореф. дис... канд. биол. наук. М., 1988. 20 с.
Перерва В.И. Особенности экологии зубра при содержании в питомниках // Биологические аспекты охраны редких животных. М., 1981. С. 33-36.
Перерва В.И. Разработка программы интродукции зубров в природе с учетом их адаптации к климатическим условиям // Научные основы охраны и рационального использования животного мира. М., 1982. С. 49-57.
Перерва В.И. Изменение экологии зубров под влиянием условий разведения в неволе // Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983. С. 151-l52.
Пуцек З. Европейский зубр: современное состояние и проблемы // Материалы I Междунар. конгр, по биосферным заповедникам (26 сентября - 2 октября 1983). Юнеско. 1986. С. 32-35.
Усов С.А. Зубр: Монография. М., 1888. 114 с.
Brincken J. Memoire Descriptif sur la Foret Imperiale de Bialowieskijen Lithuanie. N. Gucksberg. 1826, Varsovie. 1-127.
Jaczeweski L. Reproduction of European bison (Bison bonasus L.) in reserves // Acta therrlogica, Warszawa, 1958. V.1, N 9. 333-376.
Kiselewa I.G. Chow zubrow Kaukasko-bialowieskich na terenie Okskigo reserwatu. - Postepy restytucji zubra. Warszawa. 1969. 241-249.
Krasinski Z., Raczynski J. The reproduction biology of European Bison, living in reserves and freedom // Acta Theriologica, Warszawa 1967. Vol. ХII, 29. 407-444.
Krasinski Z., Raczynski J.Z. Gadan nad rozmnaraniem sie zubrow w polskich reserwatach nodowlanych i na wolnosci w Puszcry Bialowieskiej. - Postepy restytucji zubra. Warszawa, 1969. 205-231.
Оlech W. Analysis of inbreeding in European bison // Acta theriol., 1987. 32. 21-31. 373-378.
Rutberg А.Т. Birth synchrony in American bison (Bison bison): response to predation or season? // J. Mamm. 1984. 65(3). 418-423.
Wroblewski K. Zubr. Puszczy Bialowieskiey. Poznan, 1927. 1-232.